Publication LSID: urn:lsid:zoobank.org:pub:01D315F1-8FA7-4D9F-BA63-F3D5752760AC
INTRODUCTION
A peculiar new micro-gastropod from Venezuela was described by Pilsbry in 1919, as Xenodiscula venezuelensis Pilsbry, 1919. It is characterized by its nautiliform shell, sculpture of flexuous radial grooves and oddly positioned laminae in the aperture. After Pilsbry’s description of this species and genus, reports are scarce. Aside from the type taxon, only one other species, X. taintori Goodrich et Van der Schalie, 1937, was described from Guatemala and later reported from Belize, Nicaragua and Southern Mexico (Goodrich & Van der Schalie 1937, Ruiz-Cruz & Naranjo-García1998, Pérez et al. 2008, Naranjo-Garcia & Fahy 2010, Dourson et al. 2018). Moreover, X. venezuelensis was also found in Colombia and Peru (Wendebourg & Hausdorf 2019). Within Ecuador, the genus has not been reported yet (Correoso 2008, Breure et al. 2022).
In this paper, a new species of Xenodiscula, X. yumbo sp. nov., is described based on new material from northwestern Ecuador. This new record represents the first record of this genus from the Tumbes-Chocó-Magdalena biodiversity hotspot. Both other species are also discussed in this paper.
MATERIAL AND METHODS
COLLECTION METHODS
All specimens from the “Un Poco del Chocó” nature reserve (UPDC) were handpicked from dried leaflitter collected in 2019 by the first author as part of a more extensive ecological study (research permit: No. 007-2019-IC-FAU-DPAPCH-MA). The ecological data collected on Xenodiscula yumbo sp. nov. will be published in a subsequent paper containing all ecological data on gastropods from UPDC, after all taxa found in the reserve are properly described and revised (Roosen & Breure, forthcoming). The Helicinids of UPDC were already described in a previous paper (Roosen 2023).
No new material of X. taintori and X. venezuelensis has been collected for the current paper.
DESCRIPTION OF THE SPECIES
For the identification of the species Pilsbry (1919), Goodrich & Van der Schalie (1937), Dourson et al. (2018) and Breure et al. (2022) were used. In addition, the pictures of the type specimens of Xenodiscula taintori Goodrich et Van der Schalie, 1937 and X. venezuelensis Pilsbry, 1919 presented in this paper were used during the identification process of all other shells.
The pictures of all specimens were made with Scanning Electron Microscopes (SEM) and stereo microscopes at the institutes the specimens are deposited at the Royal Belgian Institute of Natural Sciences. Apertural details in Figs 7, 8 and 9 were drawn based on these images in Adobe Photoshop. Measurements were taken to the nearest 0.1 mm during the imaging process. The shell width (W) was measured at the widest section perpendicular to the coiling axis. All measurements are in mm. Whorls were counted to the nearest ¼ whorl following Gittenberger et al. (2004). Soft tissue is absent in all material, so only the shells are described.
Type material of the species described in this paper is deposited in the collection of the Instituto Nacional de Biodiversidad (INABIO), Quito, Ecuador. Other museum collections used: The Academy of Natural Sciences of Drexel University, Philadelphia, Pennsylvania, United States (ANSP) and the Universidad Nacional Autónoma de México, Mexico City, Mexico (CNMO, UNAM). Abbreviations: H – shell height, W – shell width, NW – number of whorls, Coll. – Collection, obs. – observation, TL – type locality.
SYSTEMATIC PART
Family: Scolodontidae Baker, 1925
Genus: XenodisculaPilsbry, 1919
Type species: Xenodiscula venezuelensis Pilsbry, 1919, by original designation.
Diagnosis. Animal unknown. Shell 1.1–1.5 mm, discoid, transparent or whitish with a sunken spire. Embryonic shell ½ whorl, without sculpture. Each preceding whorl is partially covered by the succeeding one. Sculpture of numerous flexuous lateral grooves and indistinct growth lines. Aperture crescent shaped, with one or more lamellae on the parietal wall. Some species have lamella on the palatal wall. Outer peristome simple, umbilicus wide.
Remarks. Xenodiscula was previously placed in the Sagdidae Pilsbry, 1895 by Pilsbry (1919). This makes sense as the Scolodontidae Baker, 1925 was not described yet and no typical scolodontids were known that develop teeth or other types of lamellae in their aperture. However, Wendebourg & Hausdorf (2019) placed Xenodiscula in the Scolodontidae based on its similarity to Drepanostomella Bourguignat, 1890. In addition, further research proved that some other scolodontids like Polygyratia J. E. Gray, 1847, Zilchistrophia Weyrauch, 1960 and Ridleyconcha Christensen, 2020 also develop palatal and parietal teeth or lamellae (e.g. Weyrauch 1960, Páll-Gergely & Asami 2014, Salvador & Cavallari 2020, Salvador 2021). Based on these arguments we agree Xenodiscula should be placed in the Scolodontidae.
Key to the species
Number of parietal teeth present is:
Only one parietal tooth . . . . . . . . . . . . . . . . . . 2
Three or more parietal teeth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . X. venezuelensis Pilsbry, 1919
Shell has:
A wide aperture with a small palatal tooth in the upper left corner. Umbilicus around 25% of total width . . . . . . . . . . . . . . . X. yumbo sp. nov.
A slimmer aperture, in which the prominent parietal tooth is the only tooth present. Umbilicus around of total width . . . . . . . . . . . . . . . . . . X. taintori Goodrich et Van der Schalie, 1937
Xenodiscula taintoriGoodrich et Van der Schalie, 1937
Figs 1–3
Species of Xenodiscula Pilsbry, 1919: 1 – Xenodiscula taintori Goodrich et Van der Schalie, 1937 (paratype, ANSP177259), Guatemala, Petén, Limestone outcrops 1 mile north-west of Paso Caballo; 2 – Xenodiscula venezuelensis Pilsbry, 1919 (syntype, ANSP105209), Venezuela, Cariaquita, leg. Brown (14.03.1911); 3 – Xenodiscula yumbo sp. nov. (holotype, INABIOEC-MCL-1214), Ecuador, Pichincha province, Un Poco del Chocó nature reserve, near Las Tolas (Gualea), coordinates (0°03'10.78″N, 78°50'32.04″W), leg. M. T. Roosen 2019. Scale bar 1 mm

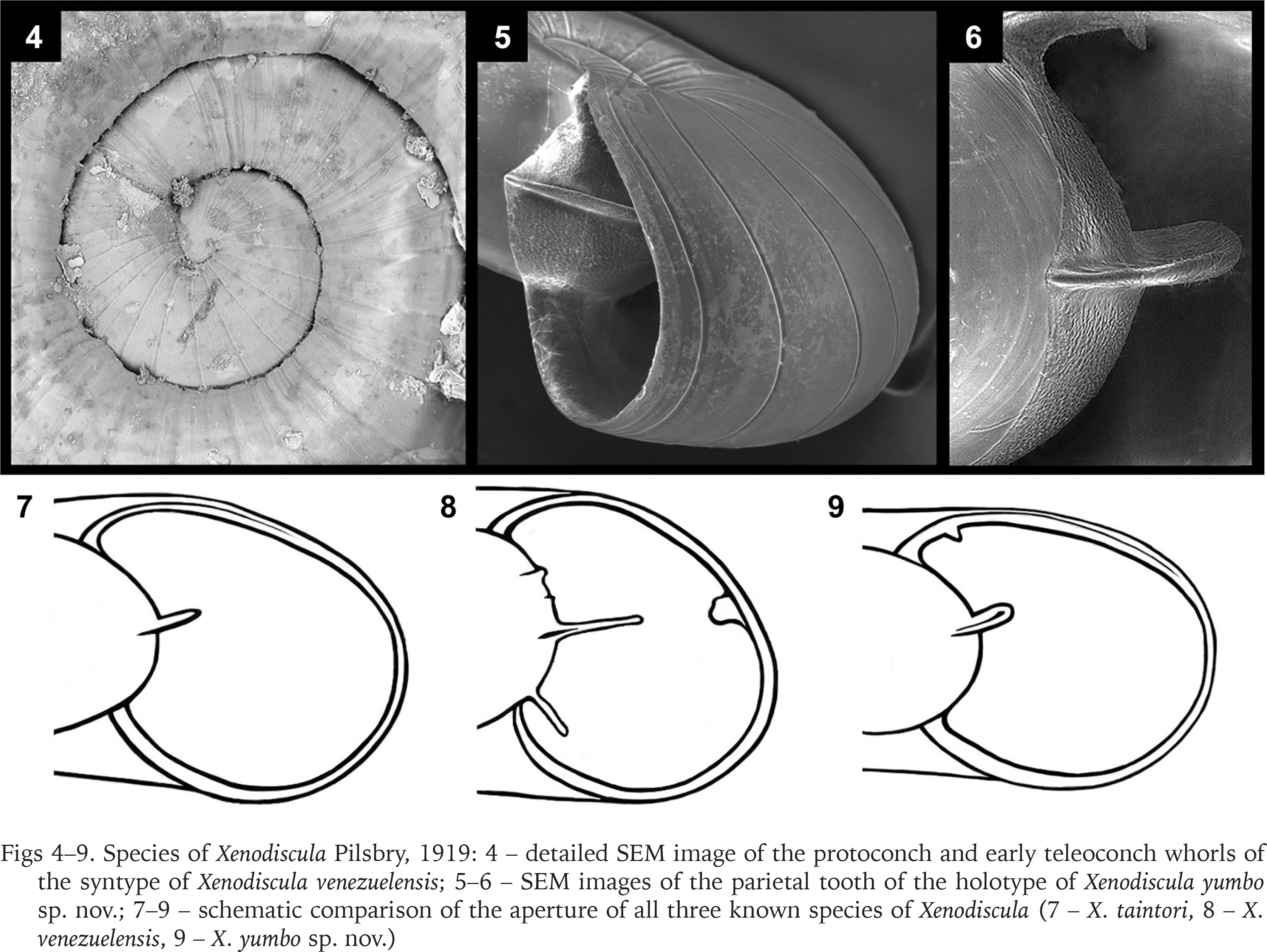
Figs 4–9
Species of Xenodiscula Pilsbry, 1919: 4 – detailed SEM image of the protoconch and early teleoconch whorls of the syntype of Xenodiscula venezuelensis; 5–6 – SEM images of the parietal tooth of the holotype of Xenodiscula yumbo sp. nov.; 7–9 – schematic comparison of the aperture of all three known species of Xenodiscula (7 – X. taintori, 8 – X. venezuelensis, 9 – X. yumbo sp. nov.)
Xenodiscula taintori n. sp. Goodrich & Van der Schalie 1937: p. 26–27, pl. 1, fig. 5a–5b.
Xenodiscula taintori Goodrich et Van der Schalie, 1937 – Dourson et al. 2018: p. 262–263.
Studied material. ANSP 177259 (paratypes, 10 shells, dry), Guatemala, Petén, Limestone outcrops 1 mile north-west of Paso Caballo. CNMO 7472 (1 shell, dry), Mexico, Chiapas, Reserva de Los Montes Azules, Río Lacanjá, leg. Lina Romero and Arturo Juárez, 24 January 2018.
Redescription. Shell small, translucent, discoid with a sunken spire. Early whorls invisible on studied pictures. Sculpture of flexuous axial grooves which are not at equal distance from each other. On the last ½ whorl, 16 grooves are visible. Whorl profile convex, aperture broadly lunulate-ovate, peristome simple. One strong, tilted upward parietal tooth at 50% of height. Parietal tooth extends past the peristome. Umbilicus wide, up to 30.8% of total width.
Dimensions. H: 0.56 mm; W: 1.3 mm; NW: 2½.
Cross-diagnosis. Xenodiscula venezuelensisPilsbry, 1919 differs by its less ovate aperture and in having more lamellae on the parietal wall. Xenodiscula yumbo sp. nov. is more similar to X. taintori, but has a more ovate aperture, less convex whorls and a small, additional tooth in the upper left corner of the aperture.
Habitat and ecology. According to Dourson et al. (2018) this species is found most often in moist leaf litter at the base of limestone hills and in moist oak forests in volcanic areas.
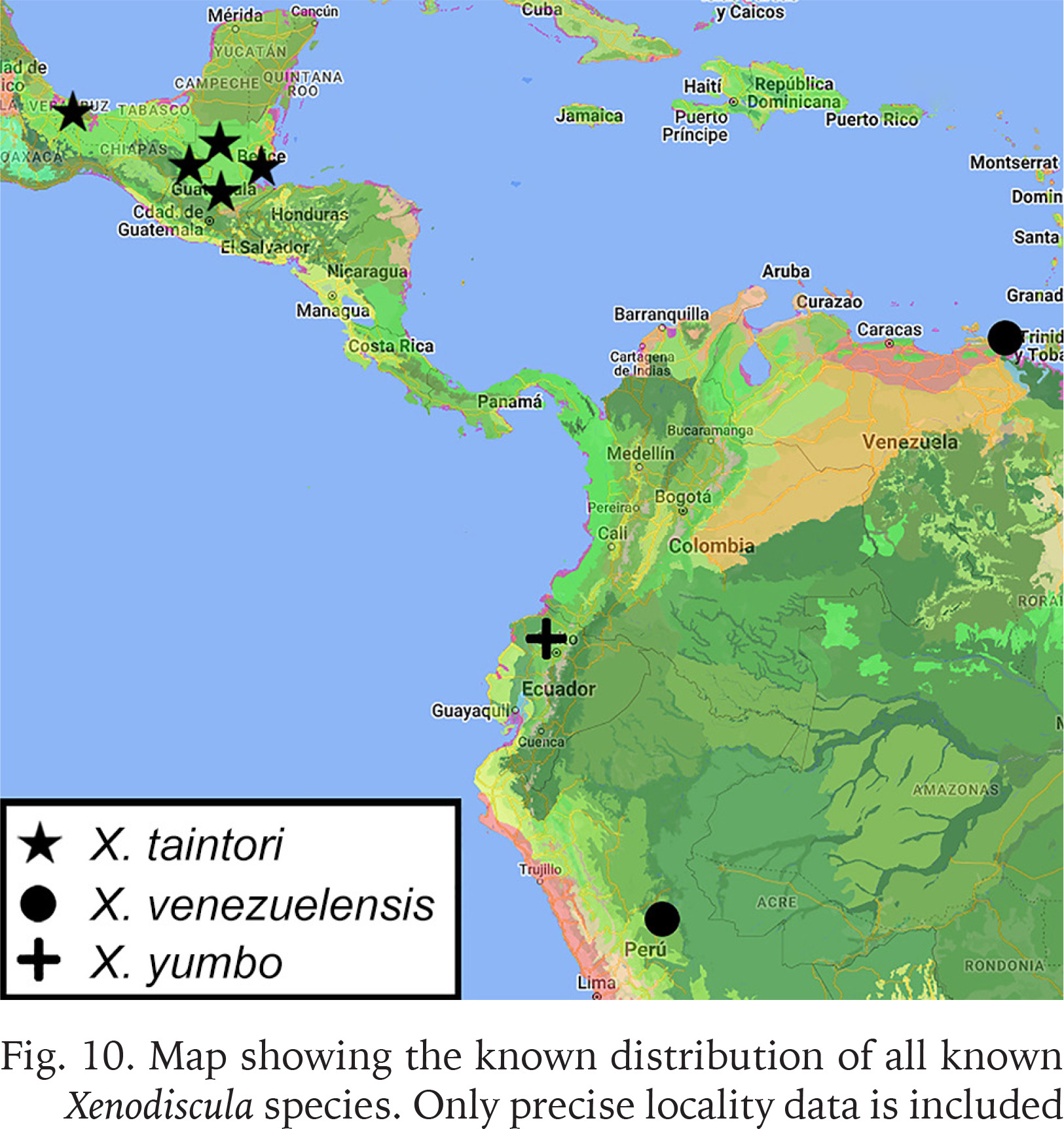
Distribution. Belize: Toledo district, base of Forest Hill, Bladen Nature Reserve (Dourson et al. 2018); Guatemala: Petén, Woodland just east of El Paso de los Caballos (TL); Alta Vera Paz (both Goodrich & Van der Schalie 1937); Nicaragua: without specific locality (Pérez et al. 2008); Mexico: Chiapas state, Reserva de Los Montes Azules, Río Lacanjá (pers. com. Naranjo-Garcia 2022); Veracruz de Ignacio de la Llave state, Los Tuxtlas biological station (Naranjo-Garcia & Fahy 2010).
Remarks. This species seems to be widespread in Central-America. However, based on images from specimens throughout its distribution, it could be a species complex. Specimens from the Bladen Nature Reserve in Belize (Dourson et al. 2018) have a much flatter whorl profile then the types. Moreover, in the sole complete specimen from Chiapas state (Mexico), the parietal tooth is less developed (pers. com. Naranjo-Garcia 2022) even though in typical X. taintori it is present at all growth stages (Goodrich & Van der Schalie 1937). We refrain from describing these forms as new species as they might represent intraspecific variation.
Xenodiscula venezuelensisPilsbry, 1919
Xenodiscula venezuelensis gen. et sp. nov. Pilsbry 1919: p. 206, fig. 1.
Studied material. ANSP 105209 (type series, 10 shells, dry), Venezuela, Cariaquita, leg. Brown (14.03.1911)
Redescription. Shell, whitish transparent, small, discoid, with a sunken spire. Early whorls partially covered by succeeding whorls. Embryonic shell ca. ½ whorl, without sculpture. Teleoconch sculpture consists of minute growth lines and irregularly spaced, slightly flexuous axial grooves which are more crowded on the oldest whorls. On the first ½ whorl of the teleoconch up to 19 grooves are present, while on younger whorls this number is reduced to 16 grooves per ½ whorl. Numerous weaker grooves are present in the spaces between grooves, aside from the shell’s growth lines. Whorl profile weakly convex, aperture broadly lunulate, peristome simple. Four lamellae are present on the parietal wall. The highest lamella is the third, which is three times as high as the fourth lamella. The upper two parietal lamellae are reduced to small ridges, while the fourth is well developed. One palatal lamella present, shaped like a rounded bulge. Last, two denticle like lamella can be seen at the base of the aperture. Umbilicus wide, up to 39.7% of total width.
Dimensions. H: 0.61 mm; W: 1.5 mm; NW: 3.
Cross-Diagnosis. Differs for X. taintori and X. yumbo sp. nov. by its less ovate aperture, larger size, more dense, stronger sculpture, relatively wider umbilicus and in having three or more lamella on the parietal wall.
Habitat and ecology.Wendebourg & Hausdorf (2019) report six specimens from leaf litter samples collected from the Panguana conservation area in the outer margins of the Amazon rainforest.
Distribution. Colombia: without specific locality (Wendebourg & Hausdorf2019); Peru: Paguana conservation area (Wendebourg & Hausdorf2019); Venezuela: Cariaquita (Pilsbry1919).
Remarks. In the original description of Pilsbry (1919), the lamellae in the aperture are described differently. First, two small lamellae were mentioned and drawn on the palatal wall, right below the main palatal lamella described in this paper. In addition, the two lamellae above the main parietal lamella are reduced or omitted in Pilsbry’s paper. Based on the description of Pilsbry (1919), we suspect that he used all 10 shells included in the type lot and described an average of them.
Because of this, it is unclear whether the specimen studied is the holotype. That said, it was taken from the type lot (ANSP 105209), so it belongs to Pilsbry’s type series. It was selected because it was an adult specimen and by that likely includes all characters that can be attributed to this species. Hopefully the redescription above will aid further research into this species.
As only SEM images of this species have been studied, it is unclear how far the lamella go into the shell.
Xenodiscula yumbo sp. nov.
urn:lsid:zoobank.org:act:B6421129-E4D9-4979-879C-17D9793AF3C9
Type material. INABIOEC-MCL-1214 (holotype, 1 shell, dry), Ecuador, Pichincha province, Un Poco del Chocó nature reserve, near Las Tolas (Gualea), coordinates (0°03'10.78″N, 78°50'32.04″W), leg. M. T. Roosen 2019; INABIOEC-MCL-1215 (paratypes, 3 shells, dry), same data as the holotype.
Type locality. Ecuador, Pichincha province, Un Poco del Chocó nature reserve, near Las Tolas (Gualea), coordinates (0°03'10.78″N, 78°50'32.04″W).
Description. Shell small, discoid with a sunken spire. Protoconch ½ whorl, unsculptured. Teleoconch sculpture consisting of flexuous axial grooves is present on the entire shell. On both the first half whorl and the last half whorl of the teleoconch 12 grooves are visible. Except for some growth disturbances, the space between the axial grooves grows exponentially correlating with the increase in whorl width. Whorl profile nearly flat, aperture broad, ovate to elongated trapezoid, peristome simple. One strong, tilted upward parietal lamella at 50% of height is present. Parietal tooth extends past the peristome. Characteristic for this species is the presence of a small palatal tooth in the upper left corner of the aperture (Fig. 9). Umbilicus wide, up to 25.5% of total width.
Dimensions. H: 0.44 mm; W: 1.1 mm; NW: 2¼.
Cross-diagnosis. From Ecuador no similar species are known. ZilchistrophiaWeyrauch, 1960 is remotely comparable in having apertural barriers, but the shape of these shells is quite different (see Páll-Gergely & Asami 2014). Moreover, Zilchistrophia does not have teeth on the columella.
The new species can be distinguished from Xenodiscula venezuelensis by its smaller size, relatively wide aperture and by having only one tooth on the parietal wall. Moreover, from apical (dorsal) view, X. venezuelensis looks less nautiliform.
Fig. 10
Map showing the known distribution of all known Xenodiscula species. Only precise locality data is included
Xenodiscula taintori is more similar to the new species by having only one parietal lamella, though it differs by its more convex shoulder, a less elongate aperture and lacking the palatal tooth in the upper left corner of the aperture. The easiest way to separate these species is by calculating the umbilical width as a percentage of the total width. Because of the elongate body whorl, the umbilicus is comparatively most narrow in X. yumbo sp. nov.
Habitat and Ecology. Fresh shells were found in leaf litter from 20 year old secondary growth Chocó rainforest at the type locality.
Distribution. Ecuador: Pichincha province, Un Poco del Chocó nature reserve (TL).
Etymology. The specific epithet refers to the Yumbo, a culture of farmers and traders who lived at the type locality until around 1660 (used as a noun in apposition). Sometimes shards of their pottery can still be found in the reserve and one of their ancient pathways is still visible on adjacent land (pers. com. Nicole Büttner 2019).
Remarks. As the specimens recovered from UPDC are smaller than or equal in size to the mesh size used to sieve the leaf litter samples, the numbers recovered are unlikely to represent the true abundance at UPDC. It is possible that the specimens found are sub-adult because of the low whorl count. According to Goodrich & Van der Schalie (1937), the parietal lamellae are already present in juvenile specimens. There are no other characters known that might help determine the growth stage of a specimen in Xenodiscula.
DISCUSSION AND CONCLUSION
In this paper the first records of Xenodiscula from the Tumbes-Chocó-Magdalena hotspot is presented. This is also the first record of the genus from Ecuador, even though a thorough study including the small scolodontids was conducted recently in a nearby area (Ramirez Perez & Hausdorf 2022).
Although this paper helps clarify the taxonomic status of Xenodiscula and provides an overview of the known species, the genus remains poorly understood. Neither soft-tissue characteristics, nor DNA or radula are known and reports are scarce. Feeding habits and ecological function are unknown. In short, there are many aspects of these species that can be studied in more detail. Hopefully this paper gives future researchers a place to start. The first step in studying these small taxa is knowing they exist and where they are found.