INTRODUCTION
Burmese amber, also known as Kachin amber or burmite, originates from Northern Myanmar and is mined in the Hukawng Valley in Kachin State. According to the uranium-lead dating (Shi et al. 2012) its age is estimated at 98.79±0.62 Ma (earliest Cenomanian). Burmese amber is very rich with fossilised organisms, with over 2770 species described or recorded so far, including both terrestrial and aquatic organisms (Ross et al. 2010, Ross 2019, 2024).
Molluscs exhibit significant diversity within Burmese amber, a stark contrast to other global amber deposits that have only sporadically yielded a few dozen land snail specimens (Poinar & Roth 1991, Roth et al. 1996, Stworzewicz & Pokryszko 2006, 2015, Balashov & Perkovsky 2020). To date 33 species of molluscs from Burmese amber have been described, as well as two more species identified to the genus-level only, with hundreds of specimens reported (Yu et al. 2018, 2021a, 2021b, 2023, Hirano et al. 2019, Bullis et al. 2020, Balashov et al. 2020, 2021, Balashov 2021, Bolotov et al. 2021, Yu & Neubauer 2021, Bichain et al. 2022, Ross 2024). Among all the molluscs found in Burmese amber, land snails represent the majority of diversity (Yu et al. 2018, 2021b, 2023, Hirano et al. 2019, Bullis et al. 2020, Balashov et al. 2020, Balashov 2021, Bichain et al. 2022). Several marine molluscs were as well reported, most notably an ammonite, Puzosia sp., gastropods, Mathilda sp., and bivalves of recently described Palaeolignopholas kachinensis Bolotov, Aksenova, Vikhrev, Konopleva, Chapurina et Kondakov 2021 (Smith & Ross 2018, Yu et al. 2019, Balashov 2021, Bolotov et al. 2021). The only freshwater mollusc found in Burmese amber, as well as globally in any amber deposit, is Galba prima Yu, Neubauer et Jochum, 2021 (Lymnaeidae) described based on two specimens (Yu et al. 2021a). Absence of other organisms embedded in the same fragments of amber has precluded definitive conclusions on how these shells of pond snails became ensnared within tree resin.
Here we report the discovery of a third specimen of Galba prima which sheds some light on the species’ ecology facilitated by the many co-occurring organisms found in the same fragment of amber.
MATERIAL AND METHODS
The amber specimen derives from the Aung Bar Amber Mine in the Hukawng Valley (vicinities of Walawbum, Kachin State, Myanmar). We are aware of the difficult situation associated with the mining of Burmese amber, involving armed conflict and civilian casualties in Myanmar since November 2017. Our sample was mined legally between October 2016 and April 2017.
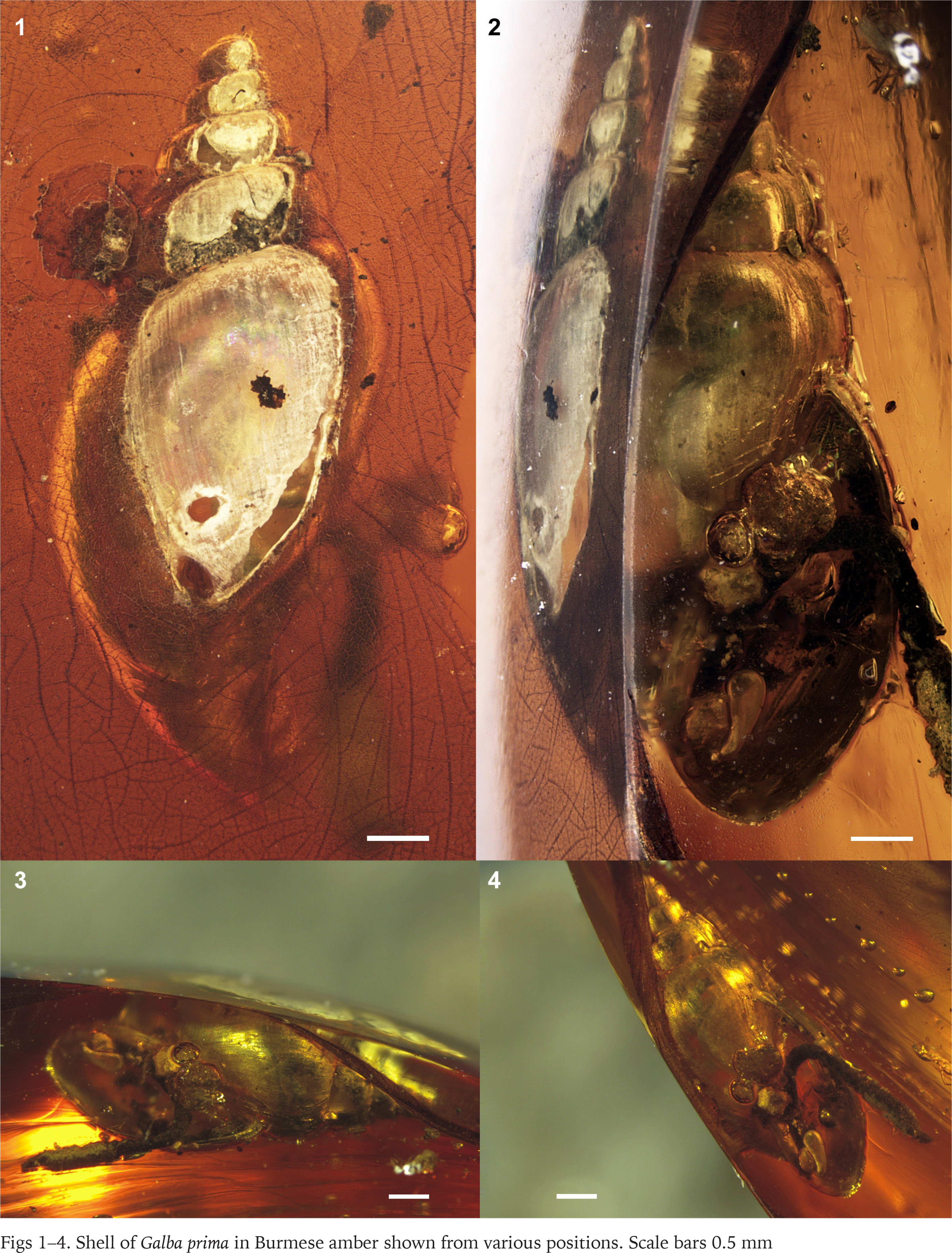
Images of the specimens were taken with Leica Z16 APO stereo-microscope equipped with Leica DFC 450 camera and processed by LAS Core software (Figs 1–2 and 5–10) and with Leica M165C stereo-microscope equipped with a digital camera and processed by Helicon Focus 6 (Figs 3–4), both at the I. I. Schmalhausen Institute of Zoology of the National Academy of Sciences of Ukraine.
Figs 5–10
Arthropods embedded together with Galba prima in the same piece of Burmese amber: 5 – Prosolierius sp. (Staphylinidae, Coleoptera); 6 – Auchenorrhyncha (Hemiptera); 7 – Nematocera (Diptera); 8 – Symphypleona (Collembola); 9-10 – various Acari. Scale bars 0.5 mm

The amber piece presently resides in the private collection of Igor Balashov, potentially at Collection of the I. I. Schmalhausen Institute of Zoology NAS Ukraine (Kyiv, Ukraine; SIZK).
SYSTEMATIC PALAEONTOLOGY
Class Gastropoda Cuvier, 1795
Order Hygrophila Férussac, 1822
Family Lymnaeidae Rafinesque, 1815
Genus Galba Schrank, 1803
Galba prima Yu, Neubauer et Jochum, 2021
Material. The shell is in a fragment of amber measuring 32 mm × 17 mm × 10 mm.
Description. Shell small, ovate-conical, consisting of approximately 6 whorls. Height : width ratio is around 1.8. Protoconch obtuse, low domed; surface details are not visible. Spire conical, apical angle approximately 65°. Body whorl large, broad and inflated, its height occupies 75% of total shell height. Whorls stepped, with a weakly convex lower part and a strongly convex upper part. Suture deep. Shell surface almost smooth, with weak growth lines. Aperture ovoid, with prominent columellar lip. Height of aperture 57% of total heigh. Peristome thin and sharp, its columellar part slightly reflected. Umbilicus small, circular, approximately 0.25 mm in diameter.
Measurements. Height of shell 6.5 mm, diameter ca. 3.5 mm; height of aperture 3.7 mm, width of aperture 2.4 mm; height of last whorl 4.9 mm.
Locality and horizon. Aung Bar Amber Mine (Hukawng Valley, Kachin, northern Myanmar); lowermost Cenomanian (mid-Cretaceous), ca. 99–98 Ma.
Syninclusions. Prosolierius sp. (Solieriinae, Staphylinidae, Coleoptera) (see Thayer et al. 2012), Auchenorrhyncha (Hemiptera), Nematocera (Diptera, at least 2 individuals), Blattoptera, Symphypleona (Collembola, at least 5), various Acari (at least 4) (Figs 5–10).
DISCUSSION
Our specimen of Galba prima exhibits striking resemblance to the holotype (Yu et al. 2021a), aligning closely with nearly all parameters and straddling dimensions between the holotype and paratype. Although it is somewhat smaller than the holotype, having about half a whorl less. Correspondingly, it represents a slightly younger specimen with near-matching proportions and size. Hence, there is no compelling basis to infer that it may represent an alternate described or undescribed species.
What is notable is that our specimen of G. prima is encased in the same piece of amber with numerous minute arthropods. At least 14 various arthropod individuals, if not more, are present in this fragment of amber. However, counting the exact number of microscopic springtails and mites is problematic due to their dual-sided visibility inside the amber, hindered by the contamination and amber structure.
All arthropods found with G. prima are terrestrial, most of them inhabiting the upper soil layer and its surface. This is consistent with the leaf litter and surface soil fauna characteristic of the tropical forest where the genesis of Burmese amber occurred (Ross et al. 2010). Notably, the abundance of soil microarthropods, including springtails and mites, clearly indicates that they were ingested in the resin in the terrestrial environment rather than as a result of resin dropping into the water. This is consistent with a soil surface of a tropical rainforest, but proximity to the shore of a small body of water remains a plausible alternative.
Considering the associated organisms, it can be assumed that our G. prima lived near a small temporary body of water located in the rainforest. This interpretation reflects the habitat preferences of modern Galba species, mostly inhabiting small freshwater bodies, including temporary ones (Økland 1990, Glöer 2019).