INTRODUCTION
Land snails constitute an essential component of global biodiversity, representing one of the most speciose groups of terrestrial animals with approximately 28,000 recognised species (MolluscaBase 2023). They are found almost everywhere on Earth (Cameron 2016) and occupy many habitat types, such as forests, shrublands, alpine meadows, urban and periurban ecosystems. The land snails of the Aegean have been studied by numerous scientists under an evolutionary, biogeographical, ecological and taxonomical framework (e.g. Fuchs & Käufel 1936, Mylonas 1982, Vardinoyannis 1994, Botsaris 1996, Welter-Schultes & Williams 1999, Cameron et al. 2000, Hausdorf & Hennig 2005, Triantis et al. 2005). Nowadays, there are more than 7,000 islands and islets in the Aegean (Triantis & Mylonas 2009), which has been vividly described as a natural laboratory of evolution, ecology and civilisations (Sfenthourakis & Triantis 2017). These maintain a hot spot of biodiversity (Myers et al. 2000) and one of Europe’s most diversified land snail faunas, with approximately 419 species (Vardinoyannis & Mylonas 2019).
The aim of the present work is a thorough study of land snail diversity in three islands of the south-eastern Aegean, namely Rhodes, Symi and Chalki. Rhodes is the largest island of the Dodecanese, with an area of 1,398 km2, 20 km off the coast of Asia Minor. The island is semi-mountainous with a maximum elevation of 1,215 m, with limestone as the main bedrock. It has a mild Mediterranean climate, and its vegetation primarily consists of coniferous forest, maquis and phrygana. Symi, located between Rhodes and Turkey, has an area of 58 km2, and its highest peak reaches 550 m (Vigla Mt.). The island consists mostly of limestone, and the vegetation is mainly coniferous forest and phrygana (Carlström 1987). Chalki is the smallest and westernmost island in this study, located about 9 km west of Rhodes, with an area of 27 km2. It is an arid, rocky island with limestones; its highest peak is Merovigli, 601 m a.s.l., and the vegetation is mainly phrygana and a few maquis. Rhodes and Chalki have been isolated from Symi and the eastern Turkish coast since the mid-Pleistocene (Lykousis 2009, Schneider et al. 2023), while Chalki separated from Rhodes during the Last Glacial Maximum (Lykousis 2009) and acquired its present configuration around 8,000 BP (Tsakiri et al. 2016). Symi was connected to the nearby mainland, the Muğla peninsula, until the beginning of the Holocene (Lykousis 2009).
MATERIAL AND METHODS
BIBLIOGRAPHIC DATA ANALYSIS
The terrestrial gastropods of some Dodecanese islands were studied in the early 20th century by Gambetta (1929) based on material collected by Festa, Desio and Ghihi-Issel. Later on, the malacofauna of Rhodes was studied by Paget (1976), Maassen (1981), Frank (1997) and Páll-Gergely & Csabai (2008) and the fauna of Chalki by Liberto et al. (2017). Until the present study, data on Symi were very scarce, usually only from sampling during the 19th century. Most of the material on which the above studies were based on was collected in late spring or summer. However, in the Aegean islands, the best period for collecting terrestrial gastropods is from late autumn to early spring, as all species are active during the wet period (Mylonas & Vardinoyannis 2022). According to previous data, 63 land snail species had been recorded from Rhodes, 14 from Symi and 22 from Chalki.
NEW DATA COLLECTION
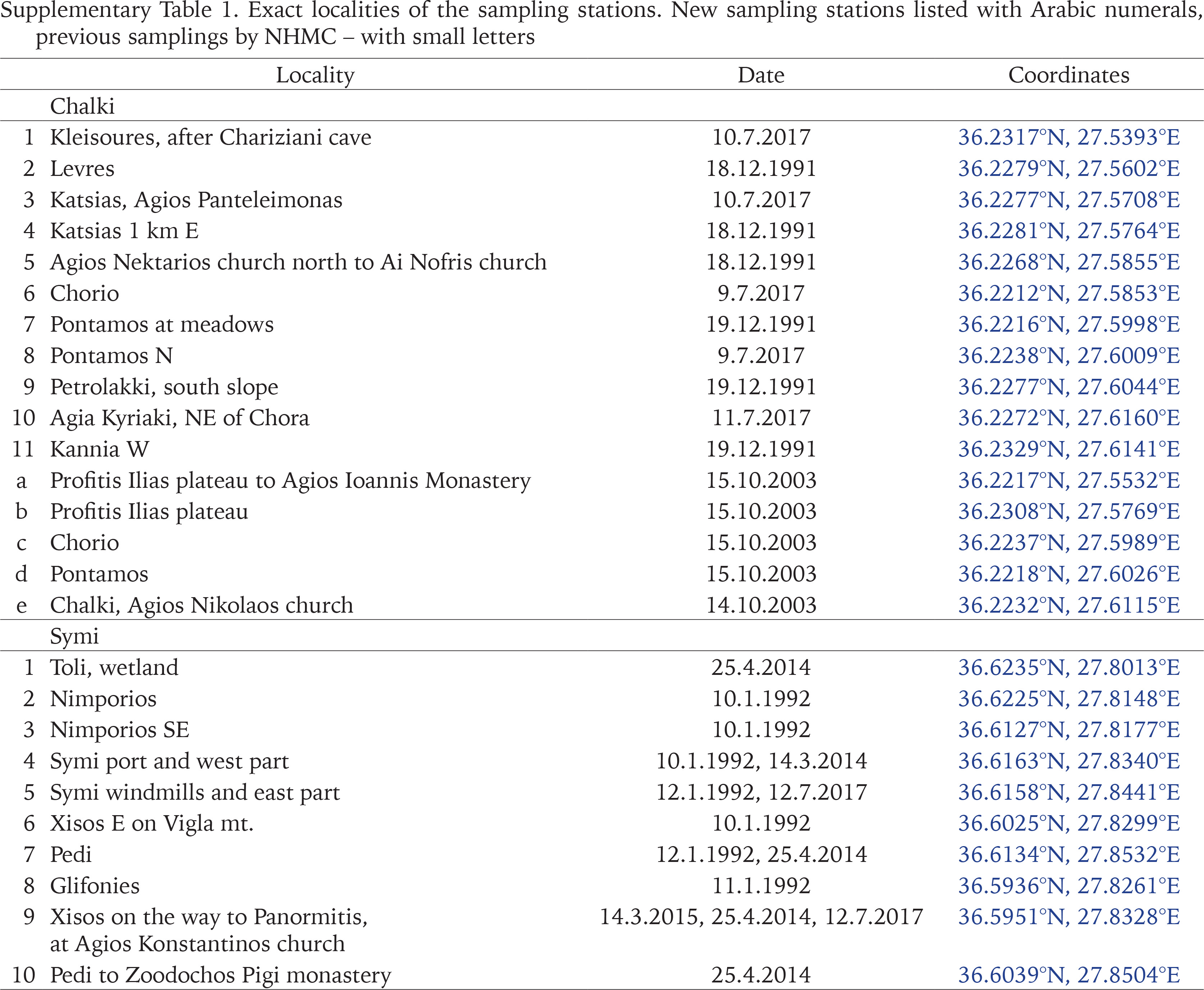
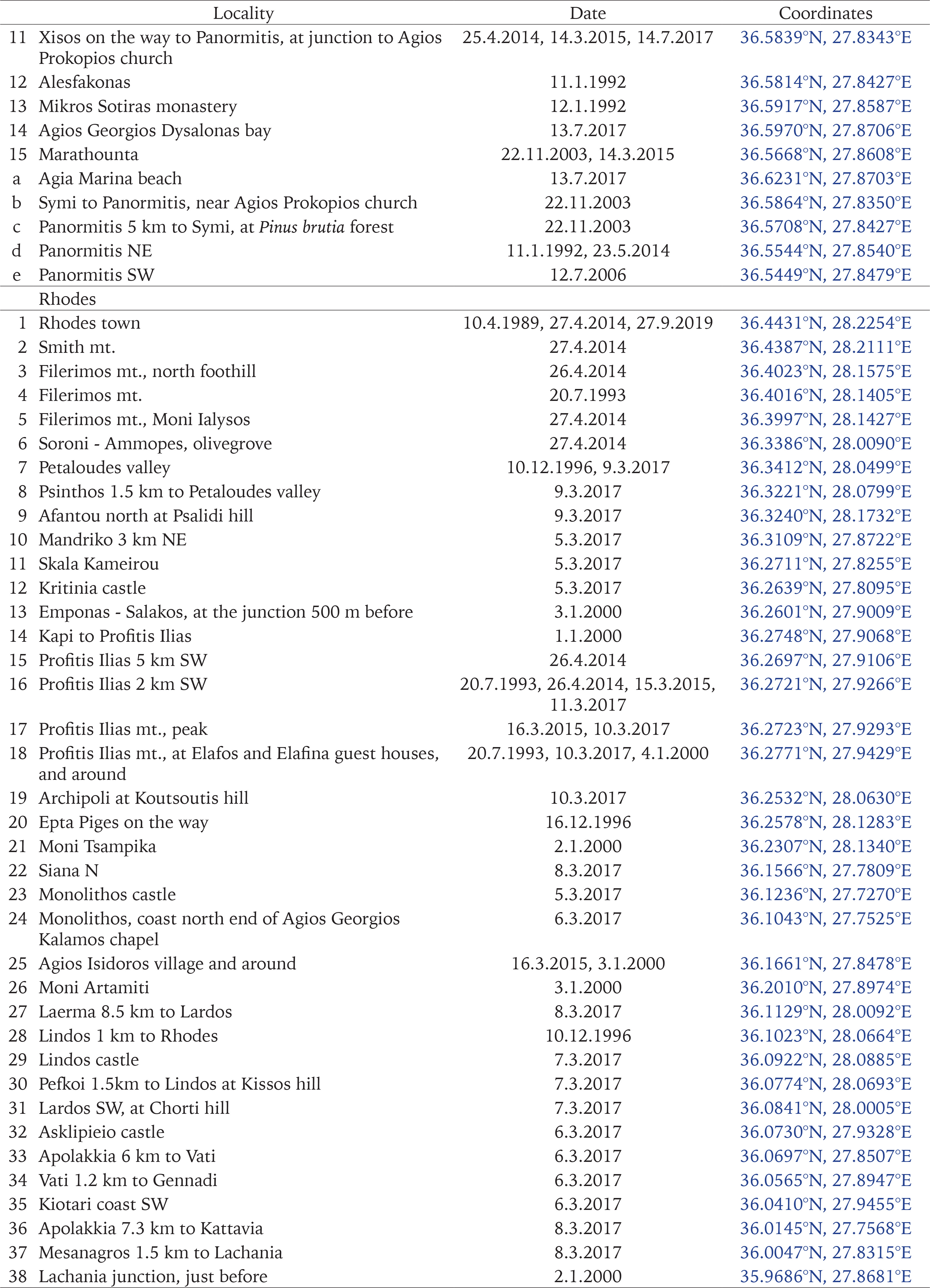
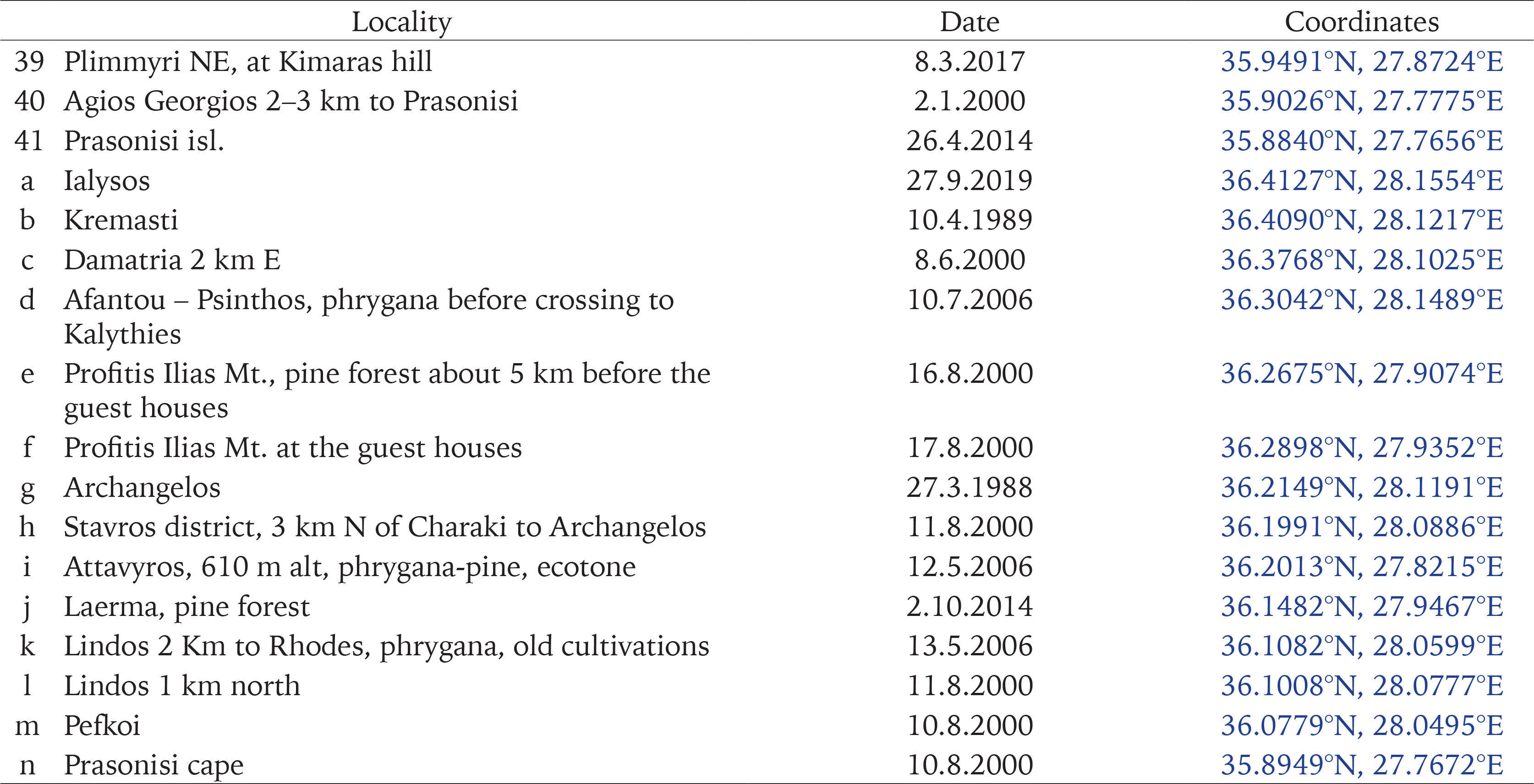
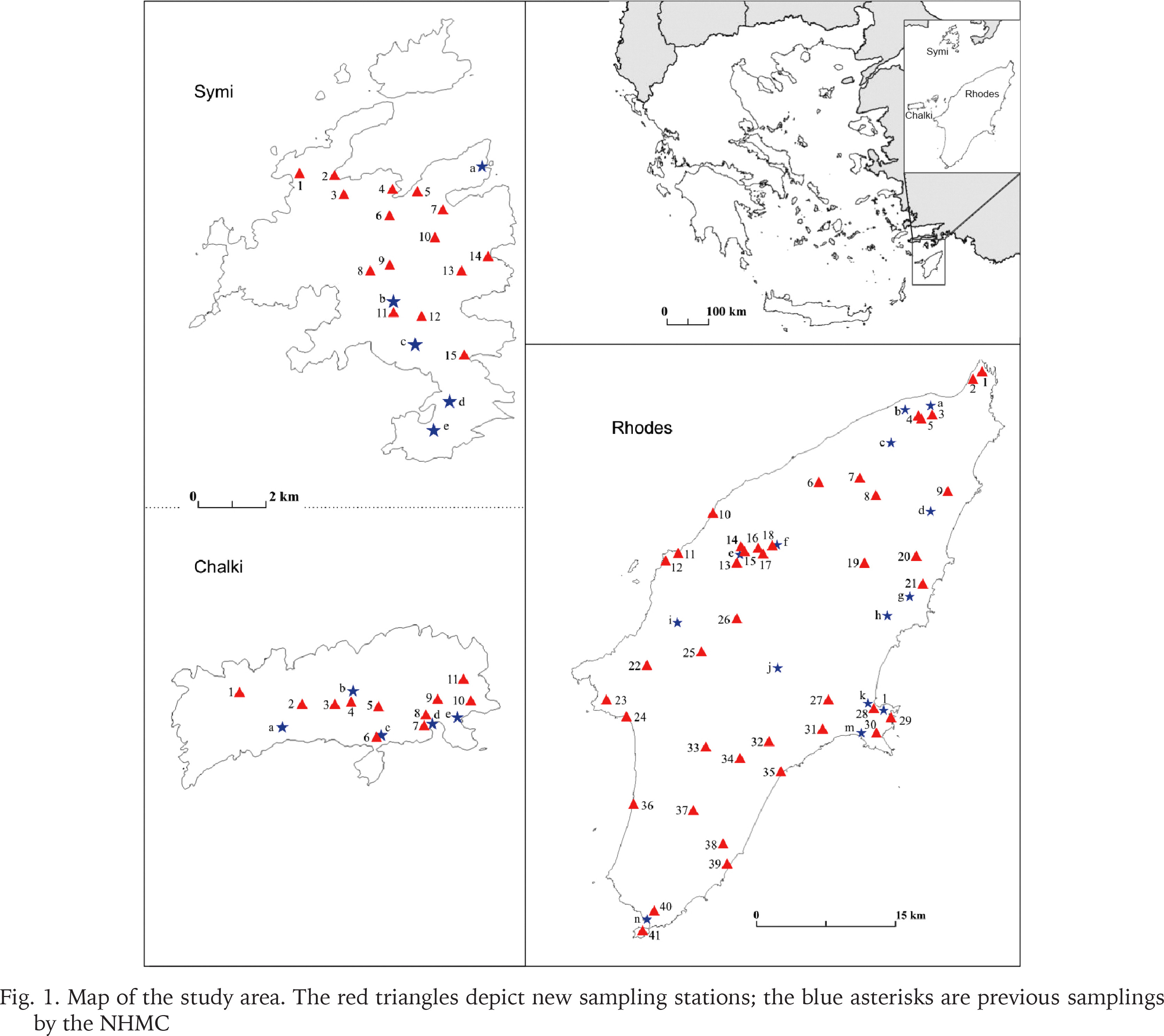
The authors collected most of the samples during several visits between 1991 and 2017. In order to collect from as many different habitats around the islands as possible, we sampled 41 sites on Rhodes, 11 on Chalki, and 15 on Symi (Fig. 1, Appendix Table S1). Snails were drowned and preserved in 75% ethanol, while specimens were also preserved in 96% ethanol for future molecular studies. Leaf litter and the upper part of the soil were collected from most sites on Rhodes and Symi, and were later sieved in the laboratory for small-sized snails. In addition, all the available material in the collections of the Natural History Museum of Crete (NHMC) was examined and used in the analyses.
Fig. 1
Map of the study area. The red triangles depict new sampling stations; the blue asterisks are previous samplings by the NHMC
Specimens were identified to the species level based on shell and anatomical characters, using the most recent scientific papers concerning each taxon (Zonitidae – Riedel 1992; Agriolimacidae, Limacidae, Milacidae – Wiktor 2001; semislugs – Schileyko 2003, Hausdorf & Kalaentzis 2021; Monacha – Hausdorf 2003; Orcullela – Hausdorf 1996, Gittenberger & Hausdorf 2004; Helix – Neubert 2014; Metafruticicola – Bank et al. 2013). All the material is held at the Natural History Museum of Crete.
It is important to emphasise that our focus is not on the subspecific level for several reasons. Firstly, many researchers struggle to determine subspecies within a species. This challenge is accentuated by the numerous discrepancies and problems with naming and identification that have been reported over the past two centuries, especially during the 19th century. However, our main concern is to clarify the terrestrial gastropod species in Greece. Having addressed this major challenge, we will then consider the potential subspecies, forms, and varieties that may be present in Greece.
RESULTS
DIVERSITY
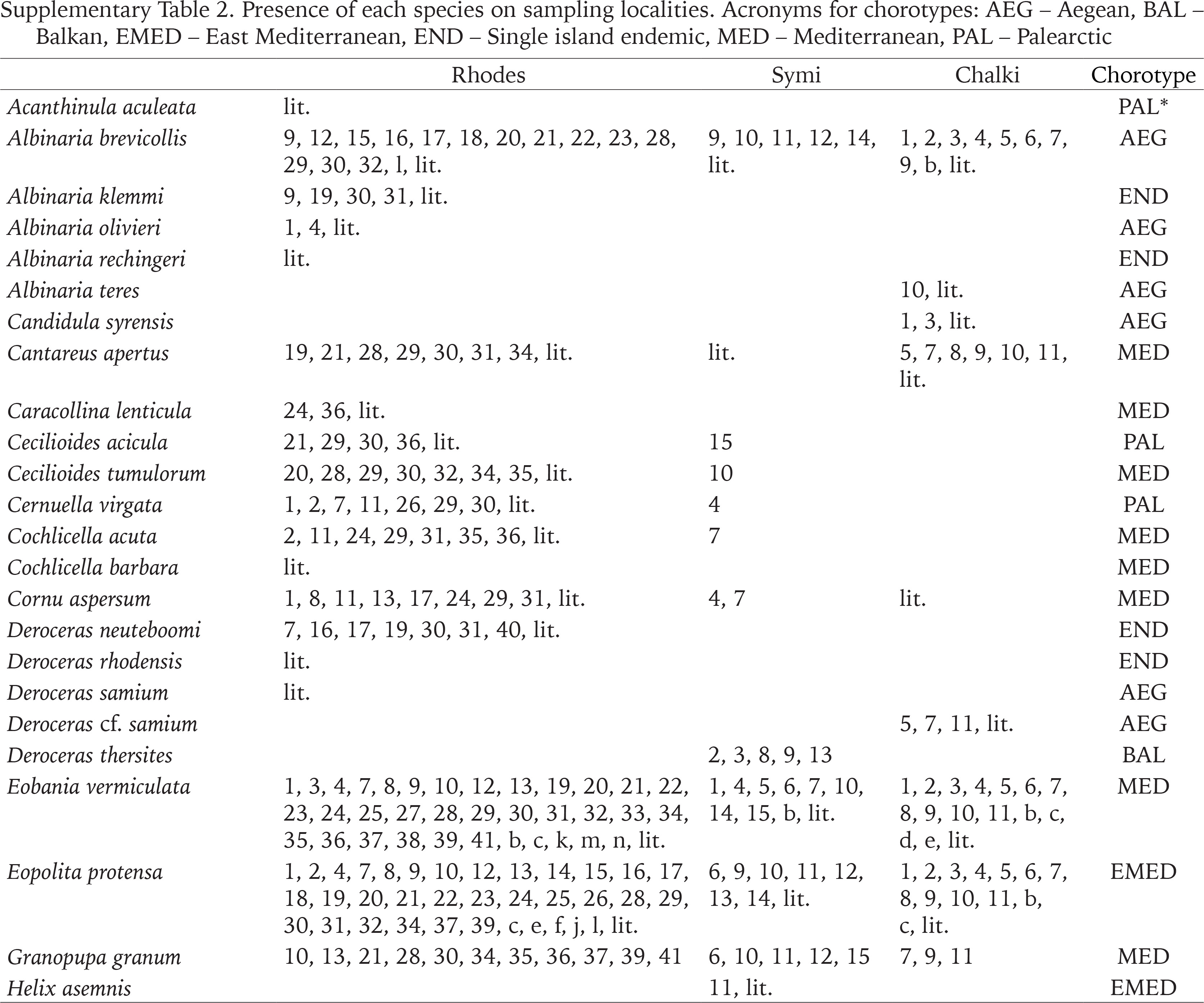
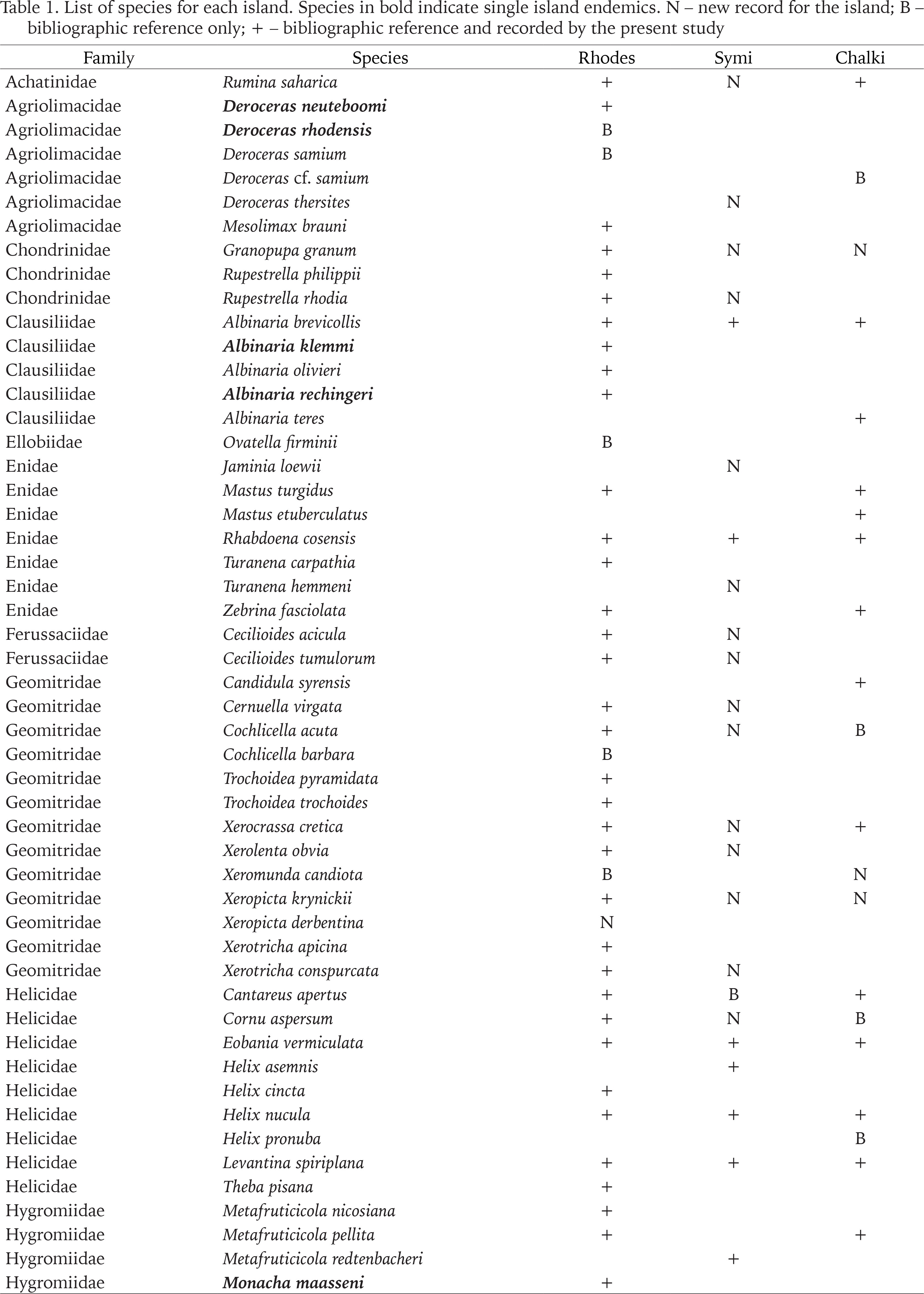
We report 67 land snail species from Rhodes, 4 of which are new records for the island, with one of them being a new record for Greece. On Symi, there are 34 species, 20 of which are recorded for the first time. Finally, on Chalki there are 32 species, with 9 new records. In total, there are 76 species present on the three islands (Table 1, Appendix Table S2), belonging to 49 genera and 25 families. The families with the most species and genera are Geomitridae (13 species, 9 genera), Helicidae (9 species, 6 genera), and Enidae (6 species, 5 genera). It is worth noting that of the 49 genera, only five are represented by more than two species, namely Albinaria (5 species), Deroceras, Helix and Vitrea (including V. sossellai (Fig. 2)) with 4 species each, and Metafruticiola (3 species), while 33 genera are represented by only one species. There are 7 single-island endemic species, all occurring on Rhodes.

TAXONOMICAL REMARKS
The first records of terrestrial gastropods in Greece date back to the beginning of the 19th century with the work of Olivier (1801). From that time until the early 20th century, taxa descriptions were mainly based on shells. Many names from this period have been clarified, either synonymised or given species or subspecies status. Those that remain unclarified or that have not been recorded since their initial record, are not included in our species list. A similar statement was made by Heller (1976) when he studied the Enidae of the Aegean.
Orculella spp.
The family Orculidae is poorly studied in Greece. In particular, the genus Orculella in the Aegean has been studied by Fuchs & Käufel (1936), Pieper (1970), Gittenberger & Hausdorf (2004), all dealing with islands in the southern Aegean. According to Hausdorf (1996) and Gittenberger & Hausdorf (2004), the taxonomy of Orculella species is based on shell characters, size, shape and sculpture, and on the genitalia, mainly the form and size of the penial appendix and of the flagellum, while Schileyko (2012) also studied the spermatophore. Based on the overall literature concerning the Aegean islands, there are 4 species of Orculella present in the Aegean islands, excluding Crete and its nearby islets. We examined numerous specimens and populations, mainly from Rhodes and compared them with specimens from other Aegean islands. Shell characters – number of whorls, lamellae and shell shape – appear to be quite plastic, even within a population, and the variation in the lamellae is extremely high. On the other hand, the characters of the reproductive system seem to be more stable. Thus, all the populations studied were classified based on their genitalia. On Rhodes and Chalki two species are present, O. critica and O. ignorata. Both species appear sympatric in only one location, Profitis Ilias 2 km SW (Rhodes). On Symi we found only O. ignorata which is the first record for the island. In certain sites that we had only empty shells, these were not assigned to either species. We must note that all older records from the Aegean islands, under the names Orcula scyphus or Orcula doliolum turcica now correspond to the genus Orculella.
Rumina saharica
Until the clarification of the Rumina taxa in the eastern Mediterranean by Bank & Gittenberger (1993), researchers had identified R. decollata and its various subspecies from Greece. However, based on the extensive Greek material in the Natural History Museum of Crete, the majority of records from Greece correspond to R. saharica. The exception is R. decollata, which is found only on a small islet off the SW coast of Crete, Elafonisos islet (KV unpublished data).
Monacha spp.
There are records of certain Monacha taxa in the Aegean islands under various names, M. cartusiana, M. olivieri, and M. parumcinta. In order to determine which taxon is present on an island, it is necessary to examine the genitalia. Since all recent studies doubt the presence of all the above species on Rhodes, and since we only found M. syriaca and M. maasseni on the island, which were identified by anatomy, we propose to remove M. cartusiana, M. olivieri, and M. parumcinta from the list of Rhodes.
Metafruticicola pellita
In their revision of the genus Metafruticicola, Bank et al. (2013) included M. pellita in the species list of Symi, based on the records of Gambetta (1929). Our thorough sampling efforts could not support this claim, since we never found it on the island, and we consider the record of Gambetta (1929) as a misidentification, therefore we exclude it from the malacofauna of Symi.
Mastus spp.
The species-rich genus Mastus is among the most common genera throughout the Aegean islands. Although there have been many studies on the systematics of the genus – by Heller (1976) (shell and genitalia), Maassen (1995) (shell, genitalia and spermatophores), Parmakelis (2003) (systematics – shell, genitalia, spermatophores, ecology, molecular) – its taxonomy is still chaotic (Bank 1997). In most of the papers dealing with the malacofauna of Rhodes or Chalki, Mastus turgidus was recorded, a name that we continue to use to avoid adding one more dubious name to islands’ fauna until a thorough study clarifies the taxonomy of the genus.
On Chalki we found M. etuberculatus and we consider that the record of Mastus sp. by Liberto et al. (2017) from this island belongs to this species, based on the data they give.
Zebrina
Liberto et al. (2017) recorded Zebrina candida (Westerlund, 1887) from Chalki as a different species from Z. fasciolata. They give a species status to the variety Bulimus fasciolatus var. β Pfeiffer, 1848, but according to Pfeiffer (1848) the variety β is unicoloured (unicolor) and white (candidus). These two Latin words have been used in the past as forms, varieties or subspecies of Z. fasciolata, e.g. B. fasciolatus forma candidus Pfr. in Westerlund (1887) or in Fuchs & Käufel (1936) as Z. fasciolata fasciolata ab. candida Pfr. Consequently, and considering that Z. candida is not a valid name in MolluscaBase (2023), we remove it not only from the faunal list of Chalki, but from the malacofauna of Greece altogether.
Additionally, Z. eburnea (Pfeiffer, 1842) was recorded from Rhodes by Westerlund (1887) under the name Buliminus calverti and also as Z. eburnea (Pfeiffer, 1842) by Heller (1976), Pieper (1984), Šešen & Schütt (2002) and Schütt (2005). However, Pollonera (1916), Bank & Menkhorst (1992) and Paget (1976) abolished this species from the list of Rhodes, which we agree with.
Chondrus zebrulus
All records of Chondrus zebrulus (A. Férussac, 1821) from Rhodes are based on a record by Westerlund (1887). However, Pollonera (1916), Paget (1976), Maassen (1981) and Frank (1997) doubted its presence on the island. Since it has never been found after its first record, in the 19th century, and most researchers doubt its presence, we remove this species from the list of Rhodes.
Deroceras cf. samium
Although the record of Deroceras cf. samium from Chalki by Liberto et al. (2017) is based on a photograph we include it in the species list of the island, but it must be confirmed by anatomical data.
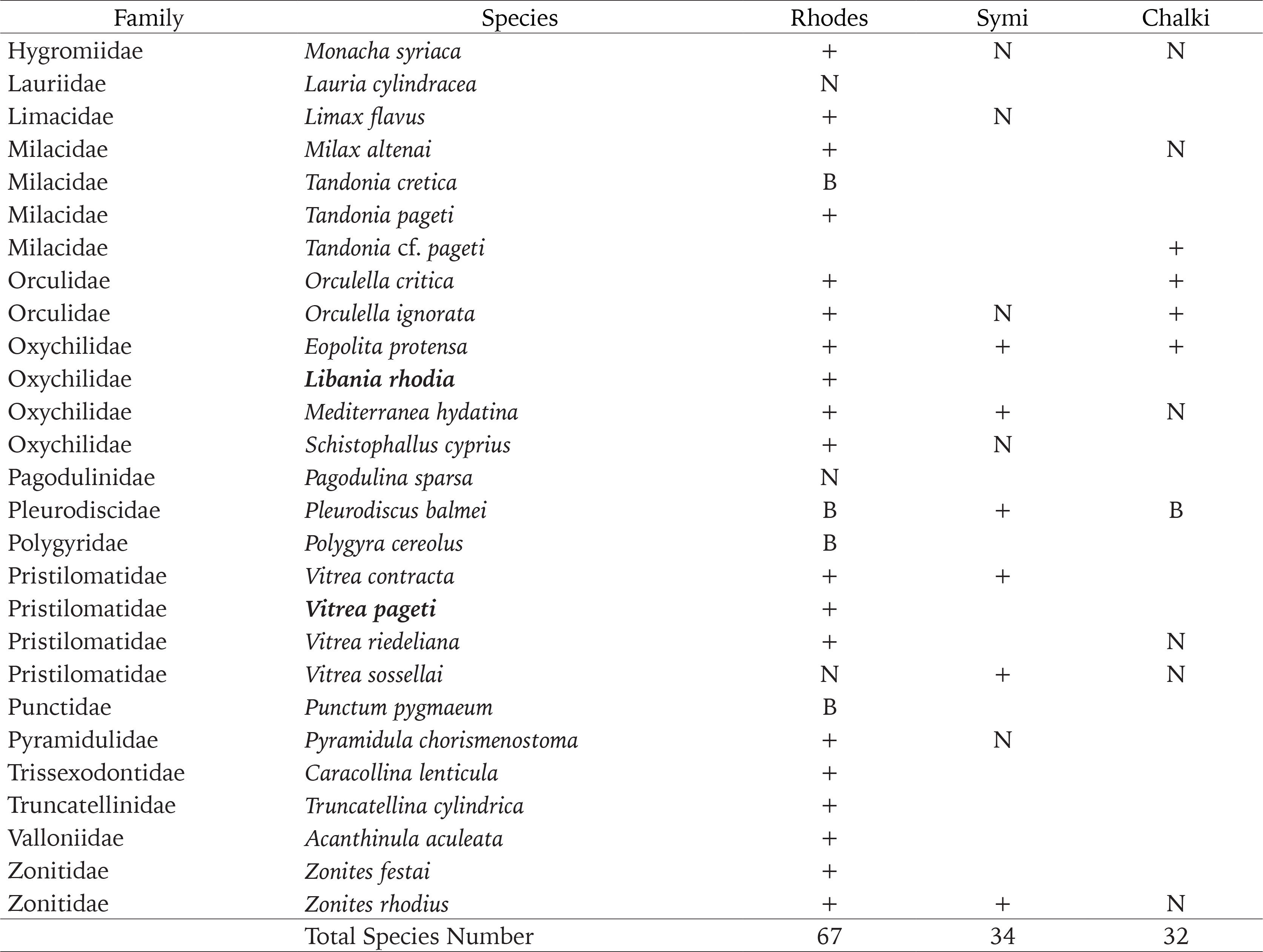
Helix cincta
The taxonomy and systematics of the genus Helix have intrigued numerous researchers (e.g. Schileyko 2006, Neubert 2014, Kórabek et al. 2015, Psonis et al. 2015). According to Neubert (2014), the most recent and undoubtedly the most detailed review of the genus in the eastern Mediterranean, the large, dark-lipped shells on Rhodes belong to H. fathallae and not to H. cincta, which constitute two closely related taxa (Neubert 2014, Kórabek et al. 2015). Neubert (2014) recorded H. fathallae from two localities on Rhodes, Lindos and Profitis Ilias Mt. 700 m altitude. Although Frank (1997) recorded only H. cincta from many sites on the island, Neubert (2014) neither confirmed nor rejected the occurrence of the latter records on the island, although on the map of the species it appears to be present on Rhodes (Neubert 2014: 91, fig. 147).
We collected ample material of dark-lipped Helix from all over the island; which gave us the opportunity to study the variation of both shell and genital characters, particularly from Lindos and Profitis Ilias Mt. peak. Shell characters, e.g. shape, size and bands, showed considerable variability (Fig. 3), whereas characters of the reproductive system were more stable (Fig. 4). In all the dissected animals, the length of the diverticulum of the gametolytic gland is less than half the length of the canal of the gametolytic gland. According to Neubert (2014: 109), the main difference between H. cincta and H. fathallae “(…) can be found in the morphology of the genital organs: H. fathallae has a well-developed diverticulum, which is missing or only rudimentarily present in H. cincta (…)”. In the same paper (Neubert 2014: 90 fig. 146, genitalia of H. cincta from Denizli, Turkey), the diverticulum of the gametolytic gland of H. cincta is well present, and its length is less than half of the canal of the gametolytic gland. Another difference in the anatomical characters of these two species is the internal features of the penis; in H. fathallae, there are “few ring-like pilasters”, while in H. cincta there are “small elongate folds” (Neubert 2014). The specimens we dissected had elongated folds and no ring-like pilasters. In addition, H. fathallae appears to be paraphyletic in the phylogenetic study of Kórabek et al. (2015), with the specimens from Rhodes to be closer to H. cincta than the Hatay populations of H. fathallae. We compared the anatomical characteristics of dark lipped Helix of Rhodes from other islands of the region, where the presence of H. cincta is not disputed (Kalymnos, Kos, Tilos), and all were found to be almost identical.
Fig. 3
Population variability of colour, shell shape and shell size of Helix cincta Rhodes isl., Afantou north at Psalidi hill (locality 9). Scale bar 20 mm

Fig. 4
Reproductive system of Helix cincta from Rhodes isl., Afantou north at Psalidi hill (locality 9). Scale bar 5 mm
Knipper (1939) and Frank (1997) questioned their respective records of H. philibinensis Rossmässler, 1839 from Rhodes. Both speculated that it might be a form of H. cincta anatolica.
In view of the above, the dark-lipped specimens of Helix from Rhodes correspond to H. cincta, and the species H. fathallae and H. philibinensis do not occur on the island.
Helix godetiana
Paget (1971) and Frank (1997) recorded H. godetiana from Rhodes based only on one very old shell. We therefore consider that Helix godetiana recorded near Lindos should not be included in Rhodes’ extant species list.
Cantareus apertus
The only record of Cantareus apertus from Symi is by Roth (1839), and it has never been recorded again. However, it may occur on the island because it can be present in gardens, cultivated fields or human-influenced habitats, being an edible and easily dispersed by humans’ snail.
Geomitridae
The family Geomitridae is remarkably species-rich in Greece. However, the taxonomy of many taxa has been based solely on shell characters, especially the ones described in the 19th and early 20th centuries. Moreover, geomitrids in Greece show a high variability in their shell characters (size, sculpture, umbilicus) (see Mylonas & Vardinoyannis 2022: fig 4A), while the genitalia seem to be a much more stable and reliable character. Thus, anatomical data of the reproductive system are crucial in resolving the taxonomy of the genera and species within the family, and one cannot base taxonomy solely on conchological characters. We decided to be cautious and not to classify geometrid specimens from sites where only empty shells were available.
Trochoidea pyramidata
A clear case of the taxonomic uncertainty in Geomitridae is Helix verticillata Pfeiffer, 1870 from Rhodes. Since its first description, based on only a few shells, this species has been cited by many authors under various generic or subgeneric names. In Tryon & Pilsbry (1888) it appears as Helix trochoides var. verticillata Parr. belonging to the section Helicella and subsection Turricula, although later Pilsbry (1894) cites it as Helicella verticillata (section Trochula Schlüter, 1838) and supports that “(…) Turricula must be rejected from the nomenclature of Helices (…)”. Gambetta (1929) gives the name Xerophila verticillata Pollonera. In web databases, it is recorded as Candidula verticillata (Fauna Europaea 2023, GBIF 2023), but in MolluscaBase (2023) as Candidula verticillata taxon inquirendum, and in the digital presentation of the Naturalis Biodiversity Center (ZMA.MOLL.401802) it appears as Trochoidea trochoides verticillata (Parr. in Pfr.). Most of the records are from Rhodes Island, with the exception of the GBIF and Naturalis Biodiversity Center records, which are from Italy. We have examined many specimens with shells corresponding to the original description of Pfeiffer (1870). There is a great variability in the shells, ranging from shells with a keel to completely rounded last whorl. Based on the genitalia, it is clear that it belongs to the genus Trochoidea due to the presence of an atrial appendix. The atrial appendix has a crest-like structure which apically is smooth (Fig. 5). This, according to Giusti et al. (1995), is characteristic of T. pyramidata and not of T. trochoides (Poiret, 1789). Thus, our specimens from Rhodes belong to T. pyramidata, and consequently H. verticillata (and all its subsequent combinations) is a synonym of T. pyramidata and should not be included in the species’ list of Rhodes.

Trochoidea trochoides
Maassen (2001) recorded T. trochoides from sand dunes in the northeastern part of the island without giving any details of the shell or the genitalia. Nevertheless, this species cannot be excluded from the species list of the island, since sand dunes are its typical habitat. Some of our sites included sand dunes but we did not find this species, however we did not collect specimens from the locality mentioned by Maassen (2001). Therefore, we include T. trochoides on the species list of Rhodes, but its presence requires anatomical verification.
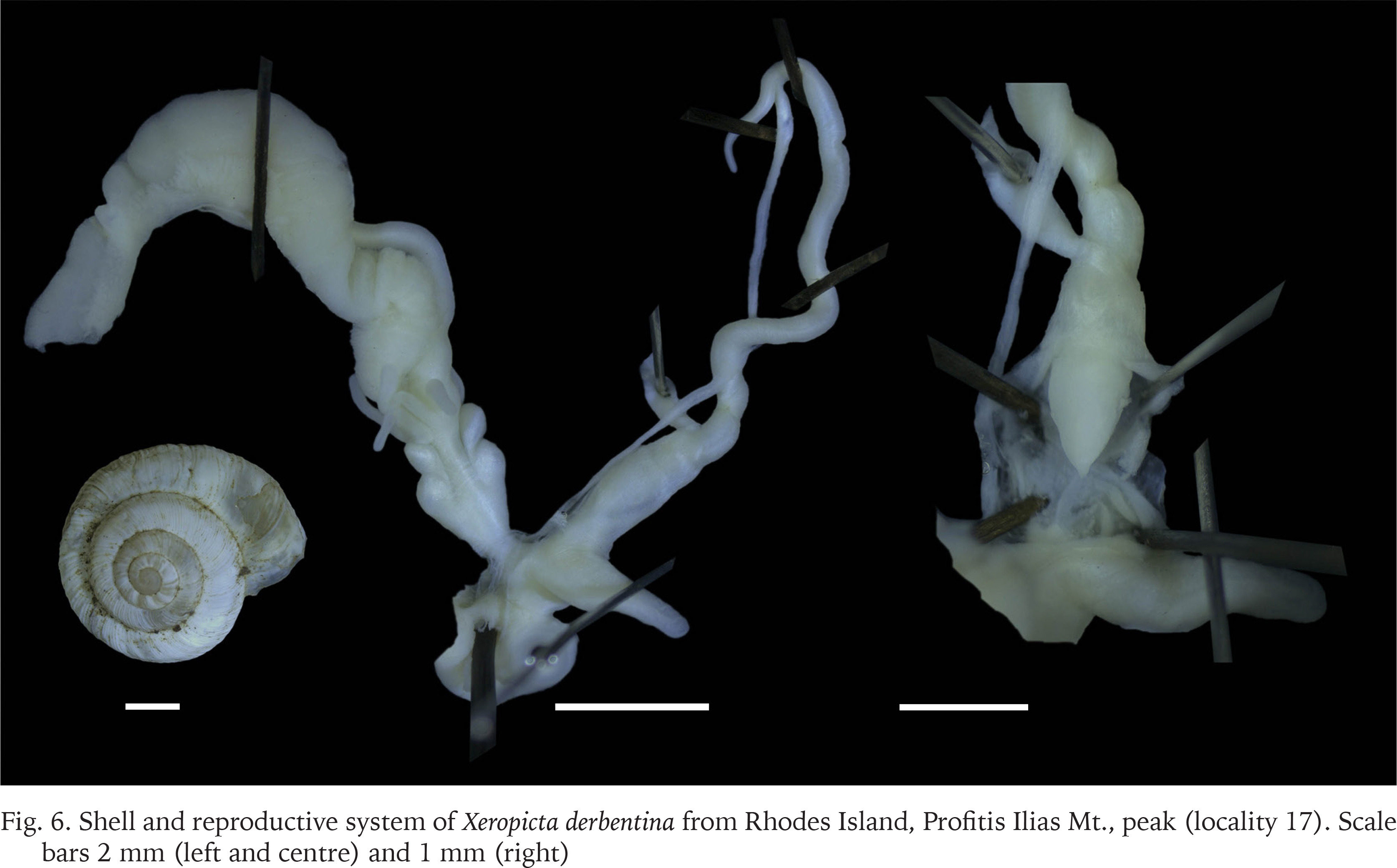
Xeropicta derbentina
Fuchs & Käufel (1936) recorded “Helicella (T.) syrensis L. Pfeiffer Rasse von Rhodes (cf. torocincta F. Haas)” from Attavyros Mt. and Profitis Ilias Mt. They claim that they have only shells, except for one specimen, and that it is similar to Xeropicta ledereri mavromoustakisi Haas, 1933 (cf. torocincta F. Haas)”. We studied many specimens (empty shells and animals in alcohol) from Profitis Ilias Mt. peak (Rhodes). Based on both the genitalia and shell characters (Fig. 6), it appears that only Xeropicta derbentina is present at the above site. The species occurs in several locations on Rhodes, and in some sites there is a high shell diversity.
Candidula syrensis
In two sites on Chalki, we found C. syrensis, a new species for the terrestrial malacofauna of the island, with a depressed and keeled shell and a very wide umbilicus. In one of the sites, we studied the genitalia, and the animals had one dart sac with a dart and a short flagellum. The specimens from Chalki were identical to those from Syros, which are deposited in the collection of the NHMC. Gambetta (1929) reported Helicella syrensis Pfr. from Rhodes, but since we did not find it on the island, we do not include it in the species’ list of Rhodes.
It should be noted that in Greece, the genus Candidula should probably be revised, since the genitalia are known from few populations of the islands or the mainland, and the named species present high shell variability. According to Chueca et al. (2018), the genus Candidula is polyphyletic; thus, new taxonomical characters should be sought to identify genera within the family Geomitridae. Clarifying the systematics of the Greek taxa requires anatomical data from many localities and a molecular phylogenetic approach, which is our future project.
In addition, the taxa Helix (Heliomanes) cornaria Berthier by Tryon (1887), Xerocrassa mesostena (Westerlund, 1879) by Martens (1889), and later Helicella (Candidula) variegata Friv. (Kattavia on Rhodes), Helicella (Candidula) hierocontina West. and Helicella (Jacosta) gradilis Pfr. by Gambetta (1929), based on samples collected by Festa, have either not been recorded again from the island or have been synonymized to species nowadays found in distant places, e.g. X. mesostena is endemic of Crete, while H. hierocontina is a synonym of Xerocrassa langloisiana (Bourguignat, 1853) which is distributed in the Levant and Egypt. Additionally, Liberto et al. (2017) recorded Xerocrassa meda (Porro, 1840) from Chalki, based on two empty fresh shells. We do not include all of the above in the species’ list of Rhodes and Chalki, respectively, because, as mentioned above, at least anatomical data are essential for the identification of geomitrid species.
DISCUSSION
Our knowledge of the terrestrial malacofauna of the three southeastern Aegean islands has increased and improved considerably. First of all, we clarified the various taxonomic inconsistencies and the different names used in the literature from the early 19th century until today. In total, 21 taxa have been clarified or eliminated from the extant terrestrial gastropod fauna of the studied islands. This has been achieved both by a thorough analysis of the published literature and by our consistent sampling efforts, especially during the wet period of the year, when the malacofauna is active and much less elusive. As a result, it is possible to find and collect well-hidden species that are aestivating deep in the ground or in rock crevices. Thus, more live specimens can be collected to study the reproductive system.
We have therefore more than doubled the number of species on Symi, added 9 species to the fauna of Chalki and 4 species to the list of Rhodes. We also added one species to the Greek malacofauna, namely X. derbentina. In contrast, we have abolished two species from the malacofauna of Greece, namely Candidula/Trochoidea verticillata and H. fathallae (both were recorded only from Rhodes).
Among the 76 species of the three islands, only 19 occur on all three, and these are species that are widespread either in the Mediterranean or in the Aegean islands (15 and 4 species, respectively). Subsequently, 41 species are found only on one island, the majority of which, 31 species, on Rhodes. The seven single-island endemic species occur only on Rhodes, the largest island of the three studied islands and geologically the oldest among all the eastern Aegean islands (Fassoulas 2018). On Symi, five species – D. thersites, H. asemnis, J. loewii, M. redtenbacheri, T. hemmeni – are not present on the other two islands but are found on other eastern Aegean islands or the nearby Turkish coast, which is to be expected, as Symi separated from the opposite mainland in the early Holocene (Lykousis 2009). Furthermore, three species on Chalki, A. teres, C. syrensis and H. pronuba, are absent from Rhodes and Symi but are present on other Aegean islands. The presence of A. teres on Chalki, the only site outside Crete, is most likely due to passive dispersal by humans (Liberto et al. 2017), as it was found next to a church.
It is worth noting that all 25 families are represented on Rhodes. On the contrary, two of the 49 genera are absent from Rhodes: Candidula, which is only present on Chalki but is widespread in the central Aegean islands and the Greek mainland, and Jaminia, which is only present on Symi but it is found on many eastern Aegean islands and the nearby mainland.
Some species are quite common on the islands and can be found in most localities, such as M. pellita, E. protensa, M. turgidus, X. cretica and M. syriaca on Rhodes, and M. pellita, X. cretica, E. vermiculata and E. protensa on Chalki. On Symi, E. vermiculata, M. redtenbacheri, L. spiriplana and Z. rhodius are the most widespread snails.
On the other hand, several species have a very narrow distribution, such as P. sparsa, R. philippii and T. carpathia on Rhodes; T. hemmeni on Symi and A. teres on Chalki. T. carpathia and R. cosensis were only found in the highest parts of Rhodes. Interestingly, T. hemmeni is specifically found in mid-altitude areas on Symi (500 m a.s.l.), whereas two congeneric species are exclusively found in high altitude – T. katerinae Gittenberger, 1996 in Crete and Cyprus and T. carpathia in Rhodes and Karpathos.
Dispersal, mainly due to human activities, is reflected in various species, either older arrivals, such as E. vermiculata and C. aspersum or more recent ones, such as X. conspurcata, X. derbentina and P. cereolus.
Notably, Rhodes has fewer species than the smaller, eastern Aegean island of Samos (72 species, 749 km2) (Bank & Maassen 1998), but the latter is much closer to the opposite mainland. On the other hand, single island endemism on Rhodes reaches 10% (7/67 land snail species) – the highest of any eastern Aegean island – while Samos and Chios each have only one endemic species. Both richness and endemism reflect the isolation of Rhodes, as it was the first eastern Aegean island to be separated from the surrounding mainland since mid-Pleistocene (Lykousis 2009). On Chalki there are more species compared to other eastern Aegean islands of similar size, such as Fournoi Island (30 km2, 22 species, Bank & Maassen 1998). This could be due to the proximity of Chalki to Rhodes and the fact that it was part of Rhodes until 8,000 years BP (Tsakiri et al. 2016), or due to a possible inadequate sampling on Fournoi. The richness and endemism of the Aegean Islands highlight that vicariance, dispersal, and the long-lasting human presence on the islands, form a continuous balance that shapes the fauna of the islands.
In conclusion, a complete and accurate knowledge of the fauna of an island in Greece is highly dependent on and linked to the sampling period. As already mentioned, the best period is when snails and slugs are active, i.e. the rainy season, from late autumn to mid spring. During this period, it is possible to find live and mature specimens of almost all species, even those that live underground, e.g. Cecilioides spp. Taxonomy is more precise and the clarification of the various names that have been reported is much easier and more reliable when we have many specimens of the whole animal and systematics are not based solely on shell characteristics, which can sometimes be misleadingly variable.