Publication LSID: urn:lsid:zoobank.org:pub:068F2F06-60AC-41C1-B3B4-F7F93D1C0849
INTRODUCTION
Within Mollusca, the freshwater gastropods form a very diverse group of organisms. This group is comprised of approximately 4,000 valid nominal species, but this estimate might represent only 25% of its actual diversity (Strong et al. 2008). These snails are found on every continent, except Antarctica, and in nearly every aquatic habitat (lotic and lentic), including lakes, swamps, rivers, streams, underground aquifers and springs, among others (Strong et al. 2008, Lydeard & Cummings 2019). The truncatelloidean family Hydrobiidae Stimpson, 1865 is the most specious assemblage of freshwater gastropods, with species occupying several biotopes, from the clear waters of mountain springs to the shallow waters of estuarine environments. Miller et al. (2018) cited 906 species, ca. 80% of them narrow-range endemics. Anatomical and systematic studies of this group are challenging due to the small size of most species (typically from 1 to 5 mm, occasionally up to 15 mm) and their reduced and simplified morphology, which yields few characters for diagnosis at the gross morphological level (Kabat & Hershler 1993, Hershler & Ponder 1998).
In contrast to this extant diversity, the fossil register is not so rich, and when referring to the family Hydrobiidae, sometimes plagued by taxonomical errors (i.e., incorrect genus assignments in the vast majority of the cases due to the lack of homoplastic characters and the simplicity of shell morphology (Hershler & Ponder 1998).
The freshwater genus Islamia Radoman, 1973, with a high species richness, is present in aquatic continental ecosystems throughout the Mediterranean region and includes 50 nominal species, 4 of which are fossils (Neubauer et al. 2014, MolluscaBase 2023). Until recently, the genus was thought to occur only in the European continental area, from the Iberian Peninsula to Anatolia (Radoman 1983, Arconada & Ramos 2006, Radea 2011, Bodon & Cianfanelli 2012, Beran et al. 2016, Yildirim et al. 2017); however, it has also been recently found in northern Africa (Glöer et al. 2020, Mabrouki et al. 2021) and it is present also in some Mediterranean islands (Bodon et al. 1995, Boeters & Beckman 2007, Radea et al. 2017). It occurs mainly in springs found at low to medium elevations, presenting a high level of local endemism (Arconada & Ramos 2006, Bodon & Cianfanelli 2012, Miller 2021).
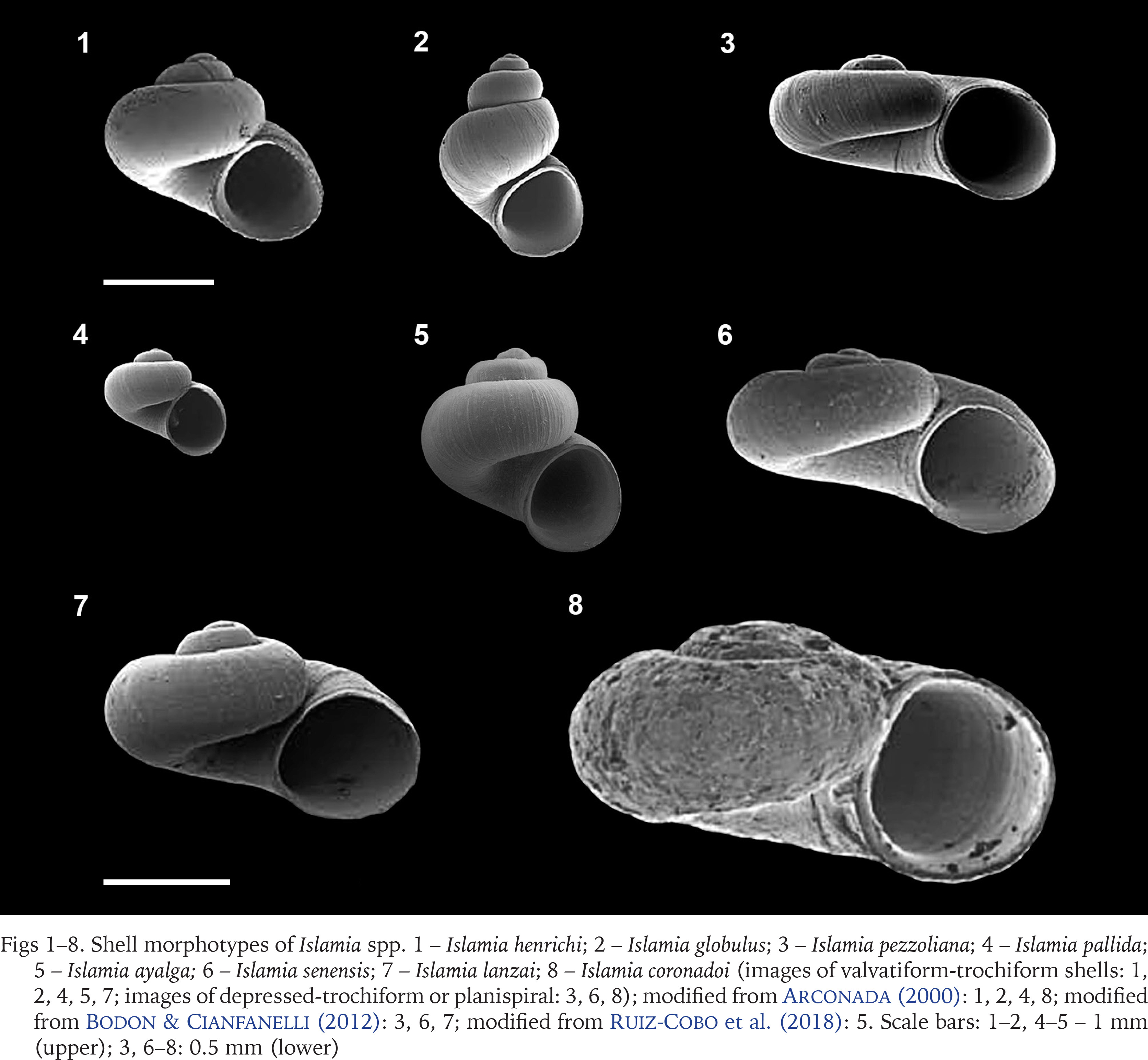
All species within the genus can be categorised into two distinct shell morphotypes based on their shape: valvatiform-trochiform shells as in Islamia henrici Arconada et Ramos, 2006 (Fig. 1); Islamia globulus (Bofill, 1909) (Fig. 2), Islamia pallida Arconada et Ramos, 2006 (Fig. 4); Islamia ayalga Ruiz-Cobo, M. R. Alonso, Quiñonero-Salgado et Rolán, 2018 (Fig. 5) and depressed-trochiform or planispiral as in Islamia pezzoliana Bodon et Cianfanelli, 2012 (Fig. 3); Islamia senensis Bodon et Cianfanelli, 2012 (Fig. 6); Islamia lanzai Bodon et Cianfanelli, 2012 (Fig. 7) or Islamia coronadoi (Bourguignat, 1870) (Fig. 8). Arconada (2000) mentioned that this character and the umbilicus width (linked to the shell form) are the only features that can help to attain a species identification based only on shell shape; otherwise, determinations depend on anatomical characters that are absent in fossil specimens. While there is a broader morphospace encompassing the range between these two extreme shapes, it contributes to enhancing the overall diversity of shell shapes within this genus.
Figs 1–8
Shell morphotypes of Islamia spp. 1 – Islamia henrichi; 2 – Islamia globulus; 3 – Islamia pezzoliana; 4 – Islamia pallida; 5 – Islamia ayalga; 6 – Islamia senensis; 7 – Islamia lanzai; 8 – Islamia coronadoi (images of valvatiform-trochiform shells: 1, 2, 4, 5, 7; images of depressed-trochiform or planispiral: 3, 6, 8); modified from Arconada (2000): 1, 2, 4, 8; modified from Bodon & Cianfanelli (2012): 3, 6, 7; modified from Ruiz-Cobo et al. (2018): 5. Scale bars: 1–2, 4–5 – 1 mm (upper); 3, 6–8: 0.5 mm (lower)
In Spain, there are just a few mentions in the literature of fossil members of this genus (e.g., Leroy 1997, De Marfà 2007, Albesa & Robles 2020). Although Albesa & Robles (2006) registered a plethora of literature between 1775–1998, there are just a few records compatible with the genus Islamia, some of them probably the result of an incorrect genus assignment as in Hauffenia schlosseri (Royo-Gómez, 1922), which Esu (1980) suggested as a member of Islamia. In the present work, a new species of the genus Islamia had been found in deposits of the Neogene in the vicinities of Alcalá de Jucar, Spain. This work aims to describe the species based on the shell shape characters mentioned before.
MATERIAL AND METHODS
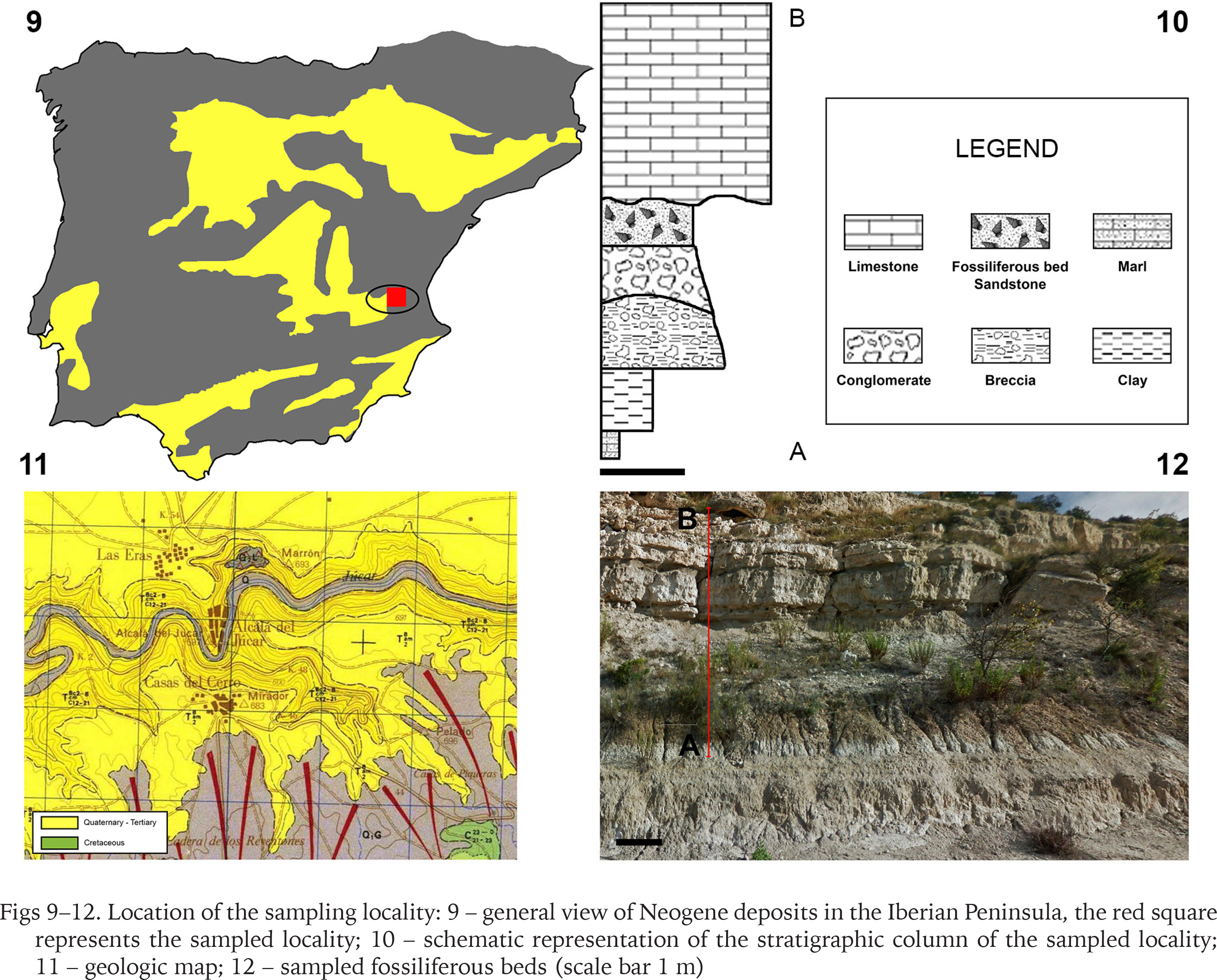
The study area according to the geological map of Spain (Alameda 1990) is composed of three main units. From bottom to top, in the first unit can be found conglomerate, calcarenite, sandstone, arkosic sand, limestone and gypsum, age Miocene (Langhian to Messinian); the second unit was composed of conglomerates, sandstones, limestone and gypsum, age Pliocene, the third unit was composed of conglomerates, limes, limestone and gypsum, age estimated Upper Pliocene to Lower Pleistocene (Figs 9, 10). Nevertheless, at a smaller scale, the sampling locality was found to contain six stratigraphic beds (Figs 10, 12). From bottom to top (Fig. 10A–B) marl, clays, breccia, conglomerates, sandstone (the fossiliferous bed) and limestone. More precise data on the stratum age were not possible to obtain as it needs more complex geochemical methods beyond the scope of this work. The estimated age for the study area was Miocene according to the geological map of Spain (Alameda 1990), but according to Esu (1980) the age of the Jucar basin was estimated from Messinian to Lower Villafranchian stages (5.6–2.59 Mya).
Figs 9–12
Location of the sampling locality: 9 – general view of Neogene deposits in the Iberian Peninsula, the red square represents the sampled locality; 10 – schematic representation of the stratigraphic column of the sampled locality; 11 – geologic map; 12 – sampled fossiliferous beds (scale bar 1 m)
The sample was composed of approx. 1 kg of sediment from the fossiliferous bed. Once in the laboratory, the sediment was dissolved in water and passed by a series of three sieves 4 mm, 2 mm and 0.5 mm mesh. In the last fraction the fossils were collected, washed with abundant running water to eliminate the limes. The fossils were then dried at room temperature and classified.
In order to obtain the shell measurements, a Leica MZ16 stereomicroscope with a Leica DFC550 camera managed from the computer by the Leica Application Suite (LAS) V4.6.13 was used. To analyse the shell microsculpture, uncoated shells were mounted on stubs and imaged on a FEI INSPECT (FEI Company, Netherlands) environmental scanning electron microscope (ESEM) at low vacuum. Maps were done using ArcGIS v.10.4 (ESRI 2011).
Abbreviations: MNCN – Museo Nacional de Ciencias Naturales; shell measurements: SL – shell length, SW – shell width, SL/SW – shell length/width ratio, BWL – body whorl length, AH – aperture height, AW – aperture width, PL – protoconch length, PW – protoconch width.
RESULTS – SYSTEMATIC
Phylum Mollusca
Class Gastropoda Cuvier, 1795
Order Caenogastropoda Cox, 1960
Family Hydrobiidae Stimpson, 1865
Genus Islamia Radoman, 1973
Islamia arcana sp. nov. †
urn:lsid:zoobank.org:act:7D47AFE7-AF18-4E87-AD4A-13E74D04DDD3
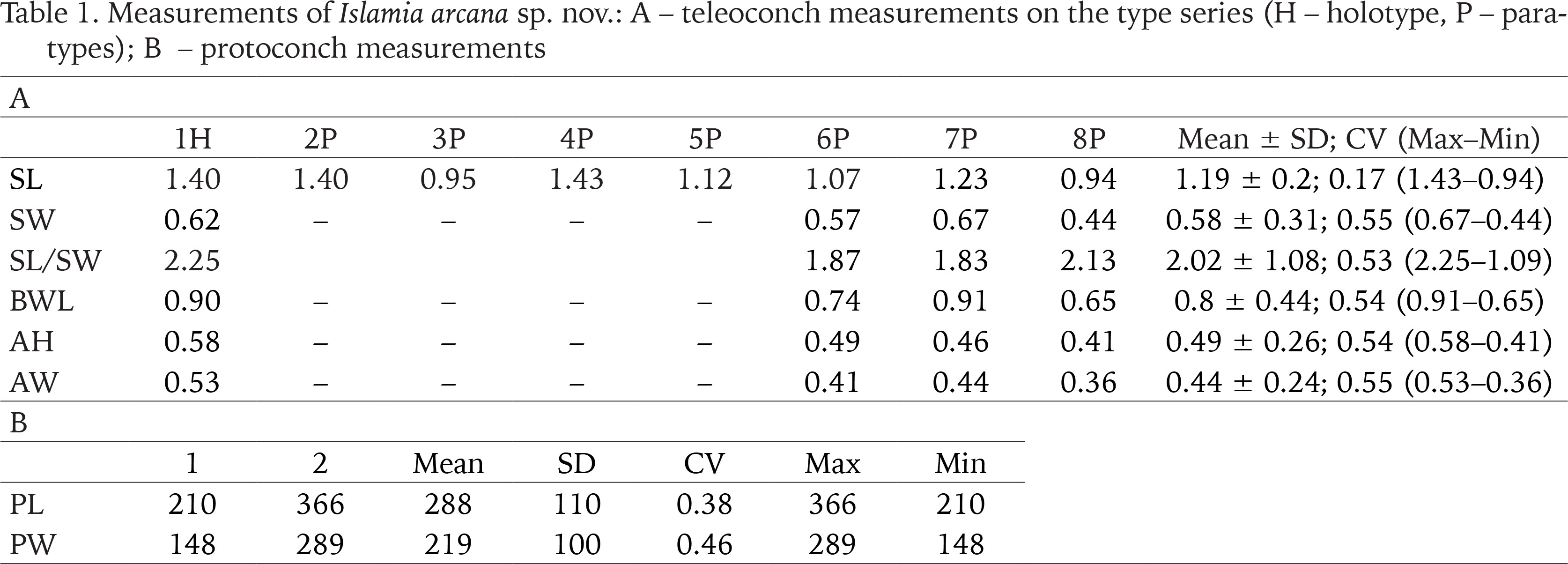
Table 1
Measurements of Islamia arcana sp. nov.: A – teleoconch measurements on the type series (H – holotype, P – paratypes); B – protoconch measurements
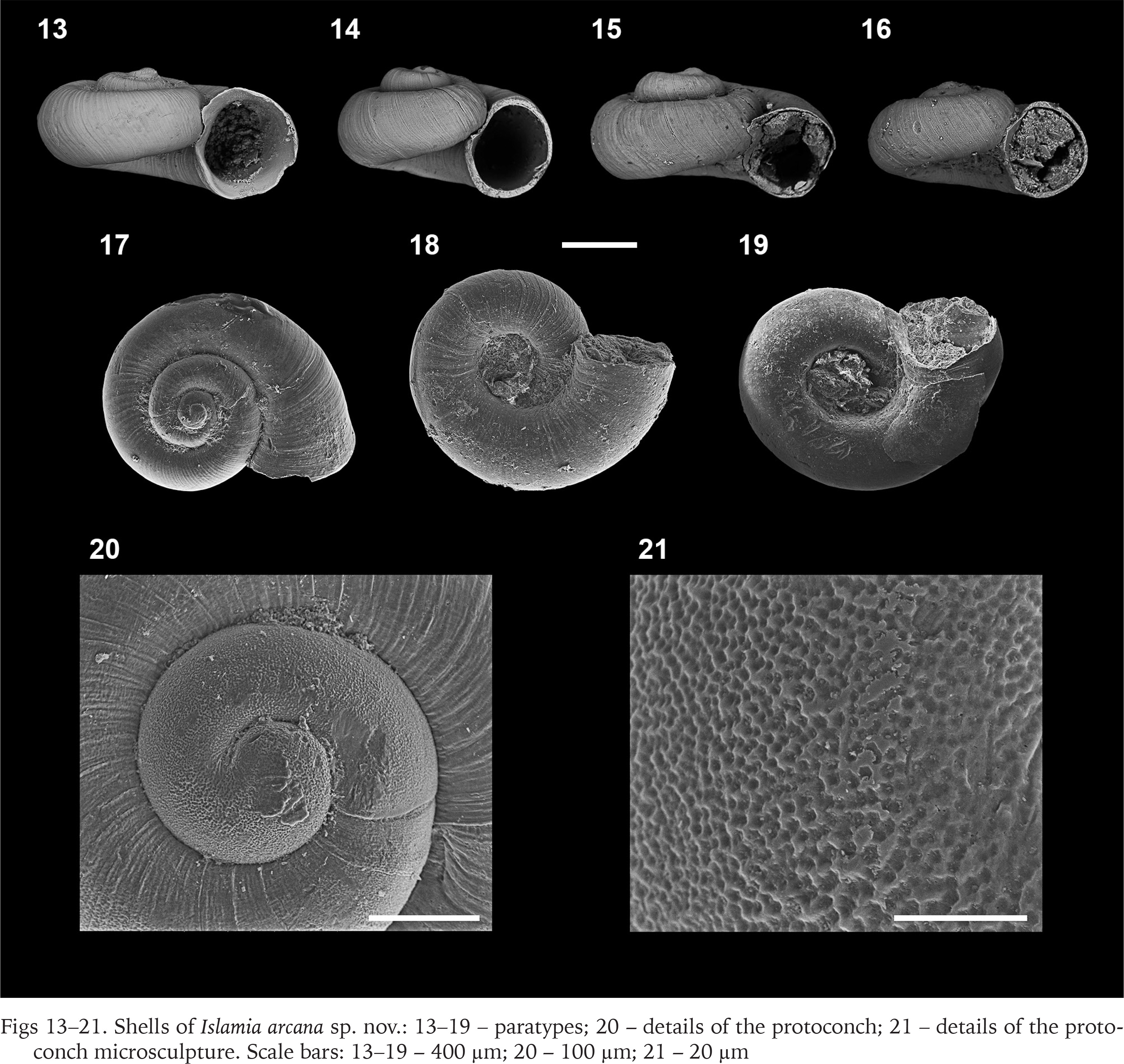
Figs 13–21
Shells of Islamia arcana sp. nov.: 13–19 – paratypes; 20 – details of the protoconch; 21 – details of the protoconch microsculpture. Scale bars: 13–19 – 400 µm; 20 – 100 µm; 21 – 20 µm
Etymology. The word arcana here refers to the Latin form arcanus, meaning hidden, secret
Type Material. Holotype: MNCNI-43550; Paratypes: MNCNI-43551 to MNCNI-43558 by present designation.
Type Locality. Road CM-3201 at the coordinates: 39.185728°N, 1.410759°W, near the town of Alcalá de Júcar, Cuenca, Spain.
Stratigraphy and age. Messinian to Lower Villafranchian stages (5.6–2.59 Mya) according to Esu (1980).
Description. Shell small; dextral, very depressed near to planispiral shape. Protoconch 210–366 µm, and one whorl, with pitted microsculpture. Teleoconch 0.94–1.19 mm; 3 whorls. Umbilicus wide; aperture rounded slightly reflexed; peristome straight; inner and outer lips about the same thickness. Shell presenting soft sinuated growth lines.
Remarks. Neogene fossil species of the genus Islamia, have been previously recorded in Spain. De Marfà (2007) cited for Fuente Nueva 3, Granada, Spain, the species Islamia minuta (Draparnaud, 1805); Leroy (1997) cited Islamia sp. for the Bañoles-Besalu Plio-Pleistocene lacustrine complex; Albesa & Robles (2020) cited Islamia sp. for Barranco León, Cuenca, Spain. Although these authors refer to the genus Islamia, it is probable that some of them refer to Valvata schlosseri (Royo-Gómez, 1922), not to a depressed planispiral valvatiform “hydrobiid”. This will be discussed further. Among the extant valvatiform hydrobiid species, Islamia arcana sp. nov. resembles Milesiana schuelei (Boeters, 1981) by its depressed shell shape, but Islamia arcana sp. nov. is larger and has more marked striation. Islamia arcana sp. nov. also resembles Islamia coronadoi (Bourguignat, 1870), but Islamia arcana sp. nov. has a wider umbilicus and a well-marked striation pattern not present in Islamia coronadoi.
In terms of shell morphology (depressed), two additional species within the genus Iberhoratia Arconada et Ramos, 2007 (in Arconada et al. 2007: fig. 2) exhibit similarities to Islamia arcana sp. nov.: Iberhoratia morenoi Arconada, Delicado et Ramos, 2007, and Iberhoratia sanromae Talaván Serna et Talaván Gómez, 2019. Islamia arcana sp. nov. displays a larger size compared to the aforementioned Iberhoratia species, and the SL/SW index value (see Table 1) is also greater in Islamia arcana sp. nov. Despite Iberhoratia sanromae presenting a striation pattern, it is less pronounced than in Islamia arcana sp. nov. The remaining species within the genus Iberhoratia feature valvatiform-trochiform shells; hence, they were not included in the comparative analysis with Islamia arcana sp. nov.
DISCUSSION
AGE ESTIMATION OF THE STUDIED AREA
The age of the sedimentary rocks of the study area had been proposed by Esu (1980), who dated the lacustrine sedimentation of the Júcar valley where this species occurs from Messinian to Lower Villafranchian stages. This corresponds to an estimated range of 5.6–2.59 Mya (Cohen & Finney 2013). The lack of a very precise age of the studied stratum is one of the weaknesses of the present study, which makes these results not applicable to other evolutionary studies that integrate molecular and fossil data to calibrate phylogenies, needed in order to understand the biogeographical history of the genus.
OVERVIEWS OF THE GENUS ISLAMIA IN THE MEDITERRANEAN SLOPE
While monotypic genera are quite common among hydrobiids, Islamia is species-rich. Distributed across the Mediterranean slope, it is a very diverse genus with 50 nominal species of which 4 are fossil species (MolluscaBase 2023). According to molecular studies, Islamia likely originated in the Italian Peninsula ca. 12 Mya, later radiating to the Balkan and Iberian peninsulas, other European territories and North Africa (Miller 2021). This agrees with the dating of I. bambolii Esu et Girotti, 2015 described for the Baccinello-Cinigiano Basin (Rook et al. 2011) dated for the Late Tortonian, Miocene (ca. 7–11 Mya). The other records of this genus belong to I. amiatae Esu et Girotti, 2015, Early Messinian, I. corinthica Esu et Girotti, 2015, Late Pleistocene. I. sarda Esu, 1984, has been re-assigned to the genus Sardohoratia Manganelli, Bodon, Cianfanelli, Talenti et Giusti, 1998.
The record of I. arcana sp. nov. constitutes the first for this genus in the Iberian Peninsula, for the Neogene. The estimated age of the Jucar Valley sedimentary rocks also agrees with the molecular clock estimated by Miller (2021) and with the proposed age of the genus. Although Islamia was cited in the literature for the Iberian Peninsula (i.e., Leroy 1997, De Marfà 2007, Albesa & Robles 2020) an analysis of the figures referred in those texts, pointed that some of the authors refer to Hauffenia schlosseri, and not to a member of the genus Islamia.
Arconada (2000) pointed out that the only two characters available to assign a given taxon to the genus Islamia were the shell shape and the size of the umbilicus, as the rest of the characters are anatomical and not applicable to the study of fossils. This author defined two primary shell morphotypes in the Islamia morphospace: valvatiform-trochiform shells and valvatiform-depressed. Fortunately, in our case, these characters have been enough as there are not plenty of extant valvatiforms with the depressed shell shape, helping to discriminate between them.
CONCLUSIONS
A revision of fossils of caenogastropods described for the Iberian Peninsula is needed in order to best characterise the palaeoenvironments and the taxonomical position of many of the described species. The vast majority were described during the last century when the family Hydrobiidae were poorly studied, and then included several taxa that today we consider as members of other families. An update is needed to review the taxonomic position assigned to those taxa at the time of its description and whether there have been subsequent changes. This task for Hydrobiidae, in general, is challenging, as mentioned at the beginning of the study. Due to morphological simplicity and high convergence in shell form, it is often difficult to elucidate genus assignments based solely on morphology. In the case of fossil forms, it is even more challenging since only conchological characters (highly homoplastic) are available, making the assignment of a specific taxon at the genus level difficult. Regarding Islamia arcana sp. nov., there was the advantage of having two distinguishing characters within the genus: the shell shape and the width of the umbilicus. These traits had been successfully applied in prior studies, demonstrating their effectiveness in identifying species within the genus. The discovery of Islamia arcana sp. nov. demonstrates that there is still work to be done in the field of taxonomy of extant hydrobioids.