INTRODUCTION
Brazil is one of the most megadiverse countries in the world for many – if not most – animal taxa (Lewinsohn et al. 2005). Nevertheless, multiple historical causes (e.g., colonialism, vast territory, inaccessible areas, lack of funding, and the current wrecking of science) have kept the knowledge of this biodiversity mostly in its infancy. That is especially true for invertebrates and, in particular, for terrestrial and freshwater gastropods (Salvador 2019, Rosa et al. 2022).
One particular region of interest in Brazil is Bahia, a large state in the east-northeastern part of the country. Despite being among the best-studied states in the country for land snails (Salvador 2019), it still hides a myriad of discoveries. For instance, that state contains the contact zones of three of Brazil’s main biomes (Atlantic Forest, Cerrado, and Caatinga) and thus, has been identified as a potential biodiversity hotspot for land and freshwater snails (Cavallari et al. 2016, Silva et al. 2021). Bahia also has numerous limestone and siliciclastic caves that have been tagged as hotspots for both terrestrial and freshwater invertebrates (Trajano & Bichuette 2009). The caves in Bahia are still largely unexplored regarding their molluscan fauna, but they should likewise harbor a wide range of potentially endemic snail species (Salvador et al. 2022b). Indeed, a few previous works have reported some gastropod species from caves in Bahia, including new species (e.g., Simone 2015, Salvador & Simone 2016, Salvador et al. 2022a), but the only more thorough molluscan survey of caves in Bahia state to date is that of Salvador et al. (2017).
Herein we examine material collected from caves in Bahia during several expeditions in the past five years. We describe a new species of Helicinidae and present new occurrences of previously known species that extend their known geographic distributions to Bahia state, including cave facultative species (i.e., troglophiles) and cave-restricted species (troglobites).
MATERIAL AND METHODS
Multiple expeditions have been conducted by the team from the LES (Laboratório de Estudos Subterrâneos, Universidade Federal de São Carlos; São Carlos, SP, Brazil) to caves in Bahia during the years 2017–2022. The collection efforts focused on a variety of animal taxa, of which the gastropods were a part. The collections were made by visual inspection of several microhabitats in the caves, using small brushes.
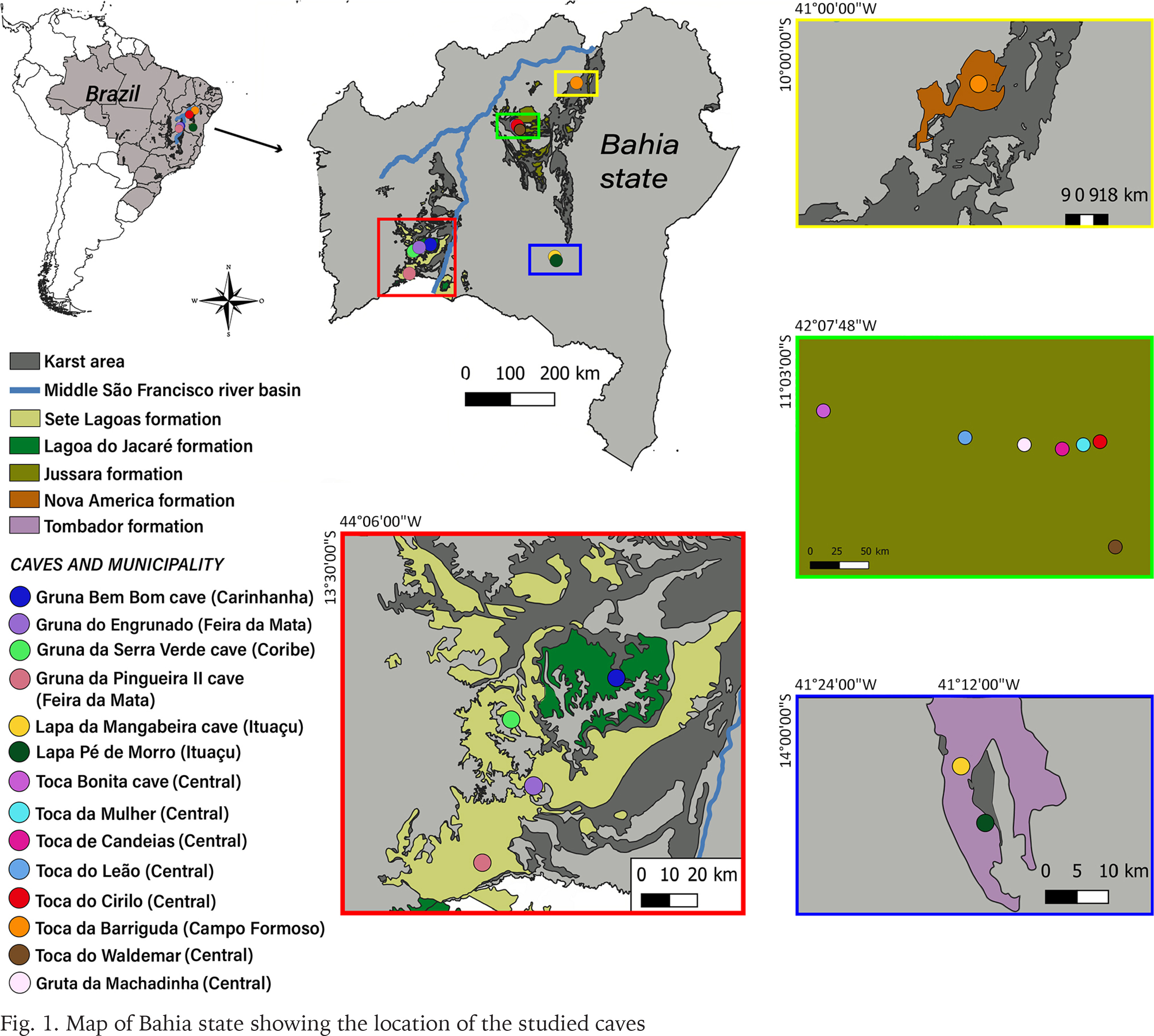
Specimens were collected in 14 of the sampled caves, which are located across the following municipalities: Campo Formoso, Carinhanha, Coribe, Feira da Mata, Central, and Ituaçu (Figs 1–4). A brief description of the municipalities’ areas and the caves can be found below.
Figs 2–4
Examples of surface and cave environments from the present study: 2 – entrance of Gruna do Engrunado cave; 3 – Gallery of Gruna do Engrunado cave; 4 – Gallery of Toca Bonita cave. Photos: Maria Elina Bichuette

The names of the caves (in Portuguese) are (Fig. 1): Toca da Barriguda (Campo Formoso municipality); Gruna Bem Bom (Carinhanha municipality); Gruta da Machadinha, Gruta do Leão, Toca Bonita, Toca da Mulher, Toca de Candeias, Toca do Cirilo, Toca do Waldemar (Central municipality); Gruna Serra Verde (Coribe municipality); Gruna da Pingueira II, Gruna do Engrunado (Feira da Mata municipality); Lapa da Mangabeira, Lapa Pé de Morro (Ituaçu municipality).
Campo Formoso municipality: Toca da Barriguda cave. This is the second largest cave in Brazil, with circa 35 km of mapped passageways (Rubbioli et al. 2019). It is a dolomitic cave of the Salitre Formation (Una geomorphological group) located in the Caatinga domain, with a tropical dry climate. The region is part of the Middle São Francisco River basin.
Serra do Ramalho karst area: Gruna Bem Bom, Gruna da Pingueira II, and Gruna Serra Verde caves. Serra do Ramalho is located in the southwestern Bahia state, on the left bank of the Middle São Francisco River basin, being dominated by a plateau formed by carbonates belonging to the Sete Lagoas Formation (Bambuí geomorphological group), with a high number of caves largely lacking legal protection (Rubbioli et al. 2019). The climate is tropical dry and the region’s main biome is the Caatinga, composed of mesophytic and xeromorphic forests interspersed with Cerrado (savannah-like vegetation) (Bichuette & Rizzato 2012).
Ituaçu municipality: Lapa da Mangabeira and Lapa Pé de Morro caves. These caves are located in the southern part of Chapada Diamantina, Upper Paraguaçu River basin. Both caves are composed of the carbonates of the Salitre Formation (Una geomorphological group; Rubbioli et al. 2019). Lapa da Mangabeira is a large cave, with more than 3 km of mapped passageways, while Lapa Pé de Morro cave reaches 1.5 km of mapped area (Grupo Araras de Espeleologia – GAE, pers. com.). The region is situated in the Caatinga domain, interspersed by rupestrian fields, with a hot semi-arid climate. The main threats to these caves come from mining activities (limestone extraction) and land use for agriculture and livestock.
Central municipality: Gruta da Machadinha, Gruta do Leão, Toca Bonita, Toca da Mulher, Toca de Candeias, Toca do Cirilo, and Toca do Waldemar caves. This region is part of the Una geomorphological group, and the caves are formed by carbonates, most of them with a few to dozens of meters of development. The climate is hot semi-arid and the region belongs to the Caatinga domain, in the Middle São Francisco River basin. The main threats to these caves are related to the installation of wind parks, altering the limestone landscapes and, consequently, impacting the caves.
TAXONOMY
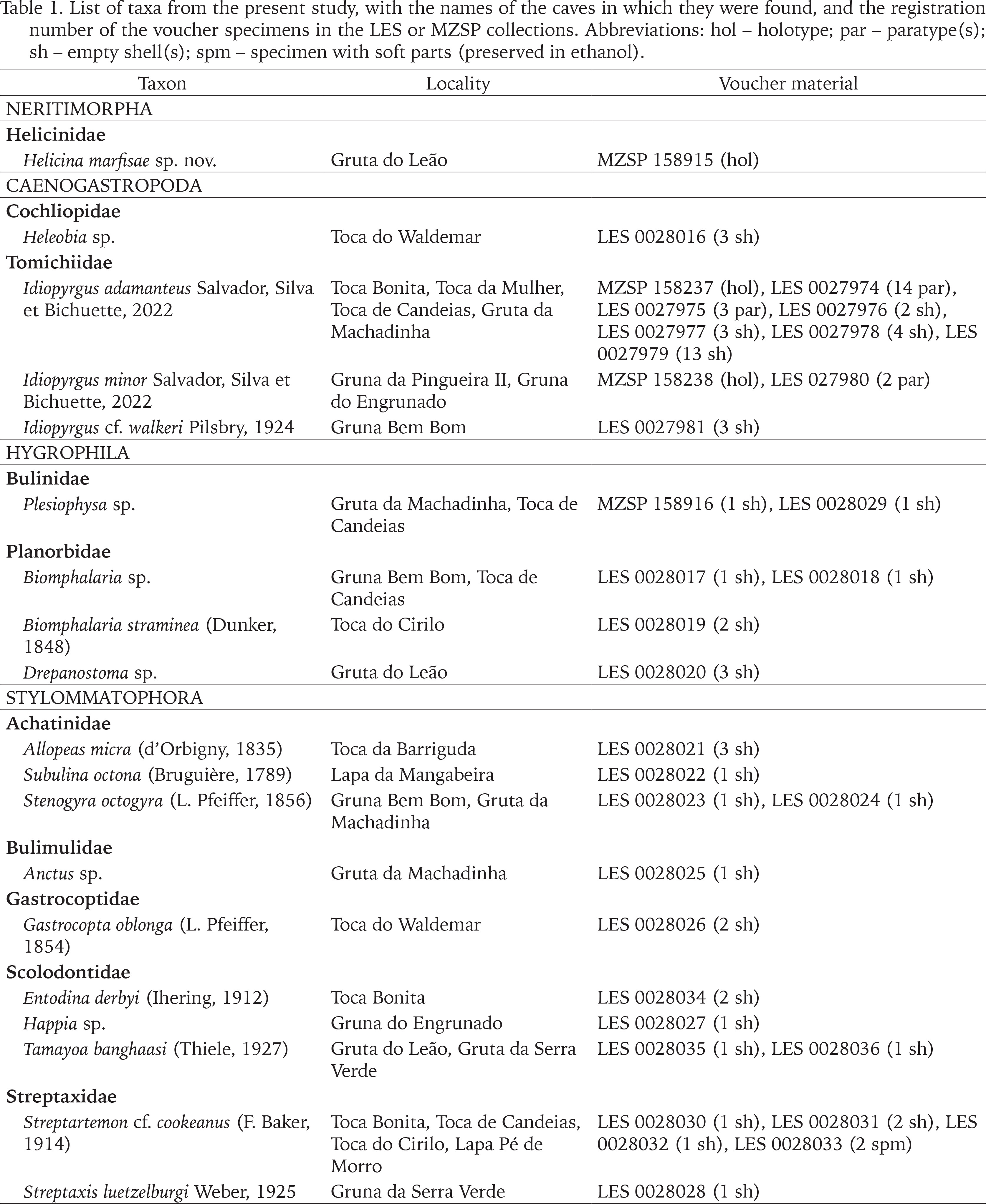
All the molluscan specimens collected in the expeditions and studied herein were deposited in the malacological collections of the Laboratório de Estudos Subterrâneos, Universidade Federal de São Carlos (LES; São Carlos, SP, Brazil) and the Museu de Zoologia da Universidade de São Paulo (MZSP; São Paulo, SP, Brazil). Empty shells were preserved dry, while complete specimens were preserved in ethanol 96%. Their registration numbers are given in Table 1. Due to conservation concerns, the precise coordinates of each cave will not be presented here; geolocation data is available in the LES and MZSP collections’ databases and can be shared upon request with the present authors or with the curatorial staff of those collections.
Table 1
List of taxa from the present study, with the names of the caves in which they were found, and the registration number of the voucher specimens in the LES or MZSP collections. Abbreviations: hol – holotype; par – paratype(s); sh – empty shell(s); spm – specimen with soft parts (preserved in ethanol).
Species identification was based on the catalogues of Simone (2006) and Birckolz et al. (2016), more specialized taxonomic works (listed in each species’ entry below, as appropriate), and comparisons with further voucher material (including types) housed in the following collections: MZSP; CMRP, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto (Ribeirão Preto, SP, Brazil); NMNZ, Museum of New Zealand Te Papa Tongarewa (Wellington, New Zealand).
Species of interest (new species and new records for Bahia state) are presented below, arranged in systematic order (Bouchet et al. 2017). A complete list of species identified in the present material (but which were already known from Bahia) is compiled in Table 1. Some specimens were juveniles or fragmentary and thus, could not be identified at species level; for those specimens, identification is possible only at the genus level. Most of the species herein are widely distributed in Brazil and consequently are not restricted to caves.
Measurements of the shells were taken with a digital caliper (0.01 mm precision, rounded to the nearest 0.1 mm) or, for minute specimens, via the software ImageJ (v.1.8.0; Schneider et al. 2012). The following abbreviations are used throughout: col. – collector(s); D – greatest shell width (perpendicular to H); H – shell height (parallel to the columellar axis); sh – dry shell(s); spm – ethanol-preserved specimen(s).
SYSTEMATICS
Subclass Neritimorpha
Superfamily Helicinoidea
Family Helicinidae Férussac, 1822
Genus Helicina Lamarck, 1799
Helicina marfisae sp. nov.
Figs 5–15
Terrestrial and freshwater gastropods found in caves in Bahia state (scale bars – 2 mm): 5–7 – Helicina marfisae sp. nov., holotype MZSP 158915; 8–9 – Idiopyrgus cf. walkeri Pilsbry, 1924, LES 0027981; 10–11 – Idiopyrgus adamanteus Salvador, Silva et Bichuette, 2022(a), holotype MZSP 158237; 12–13 – Idiopyrgus minor Salvador, Silva et Bichuette, 2022(a), holotype MZSP 158238; 14–15 – Plesiophysa sp., MZSP 158916

urn:lsid:zoobank.org:act:E4877EB4-7579-4506-930E-5F00FE7DDD13
Type material. Holotype: MZSP 158915 (1 sh; Bahia, Central, Boqueirão do Maxixe, Gruta do Leão cave, twilight zone; M. E. Bichuette, J. E. Gallão & T. Zepon col. 15/iii/2017).
Type locality. Brazil, Bahia state, Central municipality, Gruta do Leão cave, 11°03'09.0"S, 42°07'32.4"W.
Distribution. Known only from type locality. The specimens occur in the twilight and aphotic zone, in silty and rocky substrate.
Etymology. After Marfisa, a fictional character in the epics Orlando Innamorato and Orlando Furioso. Marfisa was the Queen of India and one of the fiercest warriors on the land, often compared to a lioness. The name is an allusion to the name of the type locality, which translates to Cave of the Lion.
Diagnosis. Shell minute, conical. Spire prominent in comparison to neotropical helicinids; step-like. Body whorl marked by two angulations: one on its mid-section and one on the middle of the apical area of the whorl. Aperture small, abapically positioned in relation to previous whorl.
Description. Shell minute, conical, of c. 4¼ whorls. Colour yellowish-brown, translucent; spire top and peristome white. Protoconch bulbous, of c. ¾ whorl, seemingly marked by faint malleations; transition to teleoconch marled by a sudden constriction in whorl width. Teleoconch sculpture by numerous unevenly-spaced parallel radial cordlets; cordlets absent from abapical area of whorl. Spire prominent, step-like. Suture shallow. Whorl profile convex; body whorl marked by two angulations: one on mid-section and another on the middle of apical area of the whorl. Aperture small, trapezoid, abapically positioned in relation to previous whorl; transition of columellar region to basal region of aperture marked by strong angulation; a faint angulation is present on transition of basal to palatal region. Peristome very lightly thickened, reflexed on basal and columellar regions. Shell imperforate.
Measurements. ~4¼ whorls; H – 4.4 mm; D – 4.9 mm.
Discussion. The present specimen, an empty shell, is a minute Helicinidae. Even though it is a singleton, the shell is so strikingly different from all other helicinids in the country (and in South America), that we are confident in assigning it to a new species.
All helicinids in Brazil present typical heliciniform (lenticular) shells (Simone 2006), as it would be expected; that also includes the more minute animals of similar size to the present specimen. As such, the present specimen’s minute and relatively-high spired and conical shell, with a step-like profile and abapically positioned aperture, is immediately distinct from all other known helicinids in South America. Some of its conchological traits can be observed in Central American helicinids, particularly in the Caribbean: the step-like spire is seen in Fadyenia Chitty, 1857, and the abapically-positioned aperture is seen in Lucidella Swainson, 1840. Nevertheless, the occurrence of all these traits together on a single shell is unique to the present specimen; this set of traits are thus a strong suite of morphological diagnostic characters. Thus, we consider that the present specimen belongs to a new species: Helicina marfisae sp. nov. Considering the uniqueness of its shell, it is possible that it could even represent a new genus; however, given that there is currently no phylogenetic background on Brazilian helicinids to sustain such a proposal, we provisionally classify it in Helicina.
Helicina marfisae sp. nov. can be immediately distinguished from all other helicinids in Brazil (in particular those of similar size) by its prominent step-like spire and its abapically positioned aperture. A few of the larger snails (e.g., H. concentrica L. Pfeiffer, 1849 and H. juruana Ihering, 1905, both from the Amazon) can display shells with a step-like spire profile, though their profile is much less marked than that of H. marfisae sp. nov. Furthermore, the shells of those larger Helicina spp. are at least twice the size of the shell of H. marfisae sp. nov.
The two angulations on the body whorl of Helicina marfisae sp. nov. are likewise diagnostic in comparison to other Brazilian helicinids. Many of the larger-sized species of Helicina have a mid-whorl angulation, but the presence of the more apically positioned angulation is rare, being only present on a similar scale in H. leucozonalis Ancey, 1892 from western Brazil (which is a much larger snail).
Even though the single known specimen of Helicina marfisae sp. nov. was found inside Toca do Leão cave, we cannot ascertain whether this species is troglobiotic or not, as it could also be present in unsampled areas outside the cave. The lack of obvious shell troglomorphisms (the minute size notwithstanding, as it is within the size range of the genus in Brazil; Simone 2006) could be an indication that this species is either troglophilic or even accidental, also considering it was collected in the cave’s twilight zone. Its shell, however, is rather fragile and it can be expected that it would not resist long transport into a cave, so it must be at least parautochthonous.
Subclass Caenogastropoda
Superfamily Truncatelloidea
Family Tomichiidae Wenz, 1938
Genus Idiopyrgus Pilsbry, 1911
Idiopyrgus cf. walkeri Pilsbry, 1924
Idiopyrgus cf. walkeri – Salvador et al. 2022a: p. 366.
Type locality. Brazil, Minas Gerais state, Rio das Velhas. Type material missing (see Salvador et al. 2022a).
Distribution. Known with certainty only from type locality (Salvador et al. 2022a).
Material analyzed. LES 0027981 (3 sh; Bahia, Carinhanha, Gruna Bem Bom cave; M. E. Bichuette; J. S. Gallo; J. E. Gallão col. 07/vii/2021).
Discussion. Even though the genus Idiopyrgus has been typically classified in Pomatiopsidae, a recent molecular phylogenetic study demonstrated that it belongs instead to the Gondwanan family Tomichiidae (Salvador et al. 2022a). Those authors also revised the status of all species in the genus and I. walkeri was considered a taxon inquirendum, given that its type specimens could not be located, no topotypes are known, and the simple line drawings of the species’ shell (Pilsbry 1924) are not enough to allow its identification with certainty.
Nevertheless, the wider shells of the present specimens (in comparison to other Idiopyrgus spp.) are a good match to the description of I. walkeri and its illustrations (Pilsbry 1924). As such, the specimens are here tentatively identified as Idiopyrgus cf. walkeri. Their occurrence in Carinhanha municipality is circa 450 km to the north of the river which is the type locality of the species. The specimens were in a humid substrate composed of silt and rock, in the twilight and aphotic zones of the cave.
Remarks. Two new species of Idiopyrgus were identified in the material from the present expeditions and were described by Salvador et al. (2022a) in their revision of that genus. They are I. adamanteus (Figs 10–11) and I. minor (Figs 12–13).
Subclass Heterobranchia
Superorder Hygrophila
Superfamily Lymnaeoidea
Family Bulinidae P. Fischer et Crosse, 1880
Genus Plesiophysa P. Fischer, 1883
Plesiophysa sp.
Material analyzed. MZSP 158916 (1 sh; Bahia, Central, Gruta da Machadinha cave, twilight and aphotic zones; M. E. Bichuette, J. E. Gallão & T. Zepon col. 15/iii/2017); LES 0028029 (1 sh, juvenile; Bahia, Central, Toca de Candeias cave, twilight and aphotic zones; M. E. Bichuette col. 03/vii/2018).
Discussion. Only two of the five species in the genus Plesiophysa occur in Brazil, P. guadeloupensis Mazé, 1883 and P. dolichomastix Paraense, 2002(a) (Paraense 2002a, b, 2003, Simone 2006). The latter is so far known only from its type locality in the Brazilian Midwest: a lake in Santa Rosa municipality, Goiás state (Paraense 2002a). The former, on the other hand, is widespread in Central and South America, being distributed from Guadeloupe and the Dominican Republic to southeastern Brazil (Birckolz et al. 2016). In Brazil, P. guadeloupensis has been reported from Rio Grande do Norte, Paraíba, Pernambuco, Sergipe, Bahia, Espírito Santo, Minas Gerais, and Rio de Janeiro states (Birckolz et al. 2016).
Species of Plesiophysa are considered nearly indistinguishable from one another based on shell characters alone (Paraense 2002a, b, 2003). However, the present shells are easily diagnosable from them, displaying a much more raised spired while in its congeners the spire is depressed into the later whorls. Furthermore, the shape and position of the body whorl is different in the present specimens, being shorter/smaller and much more diagonally positioned in relation to the columellar axis (i.e., at a greater angle) than in congeners. Finally, the shell aperture of the present specimens is almost perfectly oval, being more vertically positioned in relation to the shell axis.
Considering the conchological features described above, the present specimens could potentially represent a new species. However, the shells of freshwater snails are knowingly very plastic and variable in their morphology and a final verdict on the status of the present specimens will have to wait until more specimens become available, alongside anatomical and/or molecular data. For now, we identify the specimens simply as Plesiophysa sp.
Order Stylommatophora
Superfamily Streptaxoidea
Family Streptaxidae
Genus Streptartemon Kobelt, 1905
Streptartemon cf. cookeanus (F. Baker, 1914)
Figs 16–27
Stylommatophoran gastropods found in caves in Bahia state: 16–21 – Streptartemon cf. cookeanus (F. Baker, 1914) (scale bars – 2 mm; 16–18 – LES 0028031; 19–21 – LES 0028030); 22–24 – Entodina derbyi (Ihering, 1912), LES 0028034 (scale bar – 1 mm); 25–26 – Tamayoa banghaasi (Thiele, 1927), LES 0028035 (scale bar – 1 mm)

Type locality. Brazil, Rio Grande do Norte state, Ceará-Mirim and Estremoz municipalities and Baixa Verde (in Açu municipality) (Baker1914).
Distribution. Brazil: Ceará, Rio Grande do Norte and Paraíba states (Simone2006). As Salvador & Cunha (2020) argued, previous reports of S. cookeanus from southeastern Brazil belong to another species, S. waukeen Salvador et Cunha, 2020.
Material analyzed. LES 0028030 (1 sh; Bahia, Central, Toca de Candeias; M. E. Bichuette col. 03/vii/2018); LES 0028031 (2 sh; Bahia, Central, Toca Bonita; M. E. Bichuette, J. E. Gallão & T. Zepon col. 13/iii/2017); LES 0028032 (1 sh; Bahia, Central, Toca do Cirilo; M. E. Bichuette, J. E. Gallão & T. Zepon col. 14/iii/2017); LES 0028033 (2 sh; Bahia, Ituaçu, Lapa Pé do Morro; M. E. Bichuette, T. Zepon & J. E. Gallão col. 30/vi/2018).
Discussion. Shells of this species can be identified by the sloping penultimate and body whorls, alongside a proportionately small aperture that lacks teeth. The present specimens generally fit within the conchological variation observed in S. cookeanus, but some factors make a definite identification uncertain: (1) small deviations in shell morphology, such as a lower body whorl (Figs 16–18) or more laterally elongated shells (Figs 19–21); (2) the vast geographic distance to previous records (the present records would extend the species distribution around 700 km SW); and (3) the fact that the present specimens come from cave environments, which are known to foster speciation. All these could be an indication that we are potentially dealing with a new species, particularly when taking into account that specimens from southeast Brazil previously identified as S. cookeanus have been recently described as a distinct species (Salvador & Cunha 2020). As such, we consider it preferable to leave the present specimens in open classification for the time being, until anatomical and/or molecular data become available.
Superfamily Scolodontoidea
Family Scolodontidae
Genus Entodina Ancey, 1887
Entodina derbyi (Ihering, 1912)
Streptaxis (Polygyratia) derbyi Ihering 1912: p. 488, pl. 42, fig. 19.
Entodina derbyi – Morretes 1940: p. 258; Morretes 1949: p. 138; Salgado & Coelho 2003: p. 169; Simone 2006: p. 223, fig. 847.
Type locality. Brazil, Bahia state, Paraguaçu River.
Distribution. Brazil: Bahia state, São Francisco and Paraguaçu Rivers (Simone2006).
Material analyzed. LES 0028034 (3 sh; Bahia, Central, Toca Bonita; M. E. Bichuette, J. E. Gallão & T. Zepon col. 13/iii/2017).
Discussion. Entodina derbyi can be easily identified by the marked angulation on the middle section of the whorl (Ihering 1912). This species is reported from only two localities in Bahia state. The present record is thus an interesting occurrence, given that it is an intermediate point (longitudinally) between them, being closer to the São Francisco River. Furthermore, considering that E. derbyi is rarely recorded or illustrated in the literature, we hope the photographs provided here will be of help to future studies.
Remarks. The genus Entodina is often considered a synonym of Systrophia L. Pfeiffer, 1855, although sometimes it is regarded as a separate taxon (e.g., Simone 2006). Molecular data has hinted that Entodina is possibly valid as a separate genus (Salvador 2021).
Genus Tamayoa H. B. Baker, 1925
Tamayoa banghaasi (Thiele, 1927)
Happia banghaasiThiele1927: p. 319, pl. 26, fig. 13a, b, text-fig. 4; Haas1935: p. 111;
Tamayoa (Tamayops) banghaasi – Baker 1928: p. 124; Haas 1939: p. 268; Monteiro & Santos 2001: p. 1049.
Tamayoa bankhaasi – Morretes 1949: p. 139; Schade 1965: p. 218.
Tamayoa banghaasi – Jaeckel 1952: p. 7; Quintana 1982: p. 102; Oliveira & Oliveira 1984: p. 8; Santos & Monteiro 2001: tab. 2, 3; Salgado & Coelho 2003: p. 169; Simone 2006: p. 230, fig. 875; Santos et al. 2010: p. 514; Alexandre et al. 2017: p. 34; Salvador et al. 2018: p. 119, fig. 11D–G; Rangel et al. 2021: p. 4, fig. 7D.
Type locality. Brazil, Pernambuco state, Olinda municipality; and Espírito Santo state.
Distribution. Brazil: Paraíba, Pernambuco, Alagoas, Minas Gerais, Espírito Santo, and Rio de Janeiro states (Salvador et al. 2018).
Material analyzed. LES 0028035 (1 sh; Bahia, Central, Gruta do Leão; M. E. Bichuette, J. E. Gallão & T. Zepon col. 15/iii/2017); LES 0028036 (1 sh; Bahia, Coribe, Gruta da Serra Verde; M. E. Bichuette, J. E. Gallão, J. S. Gallo & V. F. Sperandei col. 05/vii/2021).
Remarks. Tamayoa banghaasi is widespread in eastern Brazil. The present records fill a gap in the species known distribution, linking the records from the northeast (Paraíba, Pernambuco and Alagoas states) to those in the southeast (Minas Gerais, Espírito Santo and Rio de Janeiro states).
DISCUSSION
A total of 19 species-level taxa (representing 10 families) was found in the present material, 8 of which are freshwater caenogastropods and Hygrophila (“basommatophorans”), while the remainder are terrestrial helicinids and stylommatophorans (Table 1). As expected, there is reduced diversity in cave environments as compared to adjacent surface areas.
The previously reported trend of Achatinidae (Subulininae) and Scolodontidae being the most diverse terrestrial families in caves (e.g., Salvador et al. 2016, 2017, 2021) is further supported by our data (Table 1). Nevertheless, it is worth noting that most representatives of those families in caves belong to widespread species in the country (Salvador et al. 2022a).
By contrast, the family Helicinidae of operculated land snails is usually uncommon among cave snail checklists, with only a few species reported from subterranean environments (Salvador et al. 2016, 2017, 2021). Nevertheless, the only new species of terrestrial snail described here, Helicina marfisae sp. nov., belongs to this family. Despite its unique shell shape (Figs 5–7), there are no obvious conchological characters that could be considered troglomorphisms (the small size notwithstanding). As mentioned above, given that only a single empty shell was found, we cannot even ascertain whether this species is indeed troglobiotic, troglophilic, or just accidental.
Among the freshwater snails, there are more cave endemics and potentially troglobiotic species (e.g., Idiopyrgus minor), likely due to the populations becoming isolated. In those cases, it is also common to find morphoanatomical features that are probably troglomorphisms, such as translucent shells and lack of body pigmentation (Figs 10–13; Salvador et al. 2022a, b). Even though Plesiophysa sp. might represent a yet undescribed species, as mentioned above, no obvious troglomorphisms can be observed in the shell (Figs 14–15).
As recently suggested in the study of Salvador et al. (2022a), it is also notable that the caves in Bahia state could be a hotbed of speciation of Idiopyrgus, an otherwise low-diversity relict genus. Besides the three species reported herein (Table 1), I. souleyetanus Pilsbry, 1911 also occurs in Bahia, totaling four out of the seven currently-recognized species in the genus (Salvador et al. 2022a).
CONCLUSION
The present study improves the knowledge of the cave snail fauna in Bahia state and has brought a new species to light. Given that the region might be a hotspot for snails and other invertebrates (Trajano & Bichuette 2009, Cavallari et al. 2016, Silva et al. 2021), the current lack of protection of the caves studied herein is worrying. The caves in the regions of Central municipality and Serra do Ramalho, for instance, are impacted by deforestation, agriculture, and mining, which increasingly shows the need for fauna studies in these places (Gallão & Bichuette 2018, Salvador et al. 2021, 2022a).