INTRODUCTION
South American megasnails are terrestrial stylommatophoran gastropods belonging to the genus Megalobulimus K. Miller, 1878 (Strophocheilidae, Megalobuliminae). They are distributed throughout South America with major areas of diversity in the Andean Amazon and eastern Brazil (Bequaert 1948, Mansur & Leme 1996, Ramírez et al. 2012, Borda & Ramirez 2014). While some species (e.g., M. oblongus (O. F. Müller, 1774)) are widely distributed across the continent, many (e.g., M. proclivis (E. von Martens, 1888) and M. cardosoi (Morretes, 1952)) have much more restricted distributions (Bequaert 1948, Leme & Indrusiak 1995, Salvador et al. 2018).
The present classification of Megalobulimus, single member of subfamily Megalobuliminae, is mostly based on shell morphology (Bequaert 1948, Morretes 1952), which has been largely supported by anatomical features (Leme 1975a). Presently, the genus includes over 80 species, circa 60 of which occur in Brazil (Simone 2006, Birckolz et al. 2016, Fontenelle et al. 2019, 2021).
Megalobulimus spp. are herbivores and typically occur in low densities, living in forest litter and soil and being active in the night or immediately after rain (Oliveira et al. 1984, Fontenelle & Miranda 2017, Miranda et al. 2015, 2020, Miranda & Correia 2021). Their large size inevitably meant that they would be used for human consumption, both for the first peoples of South America, as well as by later migrants (e.g., Martens 1876, Valduga 1985, Araujo et al. 2012, Ramírez et al. 2012). Their large and thick shell also imply a greater dependency on calcium sources (Dallinger et al. 2001). Calcium is directly related to the abundance, development, reproduction and survival of the large African Achatinidae snails (e.g., Ireland 1991, Egonmwan 2008) and a similar situation is expected with Megalobulimus. Some species seem to have an affinity for limestone soils and rocks, occurring in greater numbers close to sheltered areas such as caves (Vieira & Simone 1990, Salvador et al. 2021, J. H. Fontenelle pers. obs.). The depositional environment of such cases also favors the preservation of the shells as subfossils.
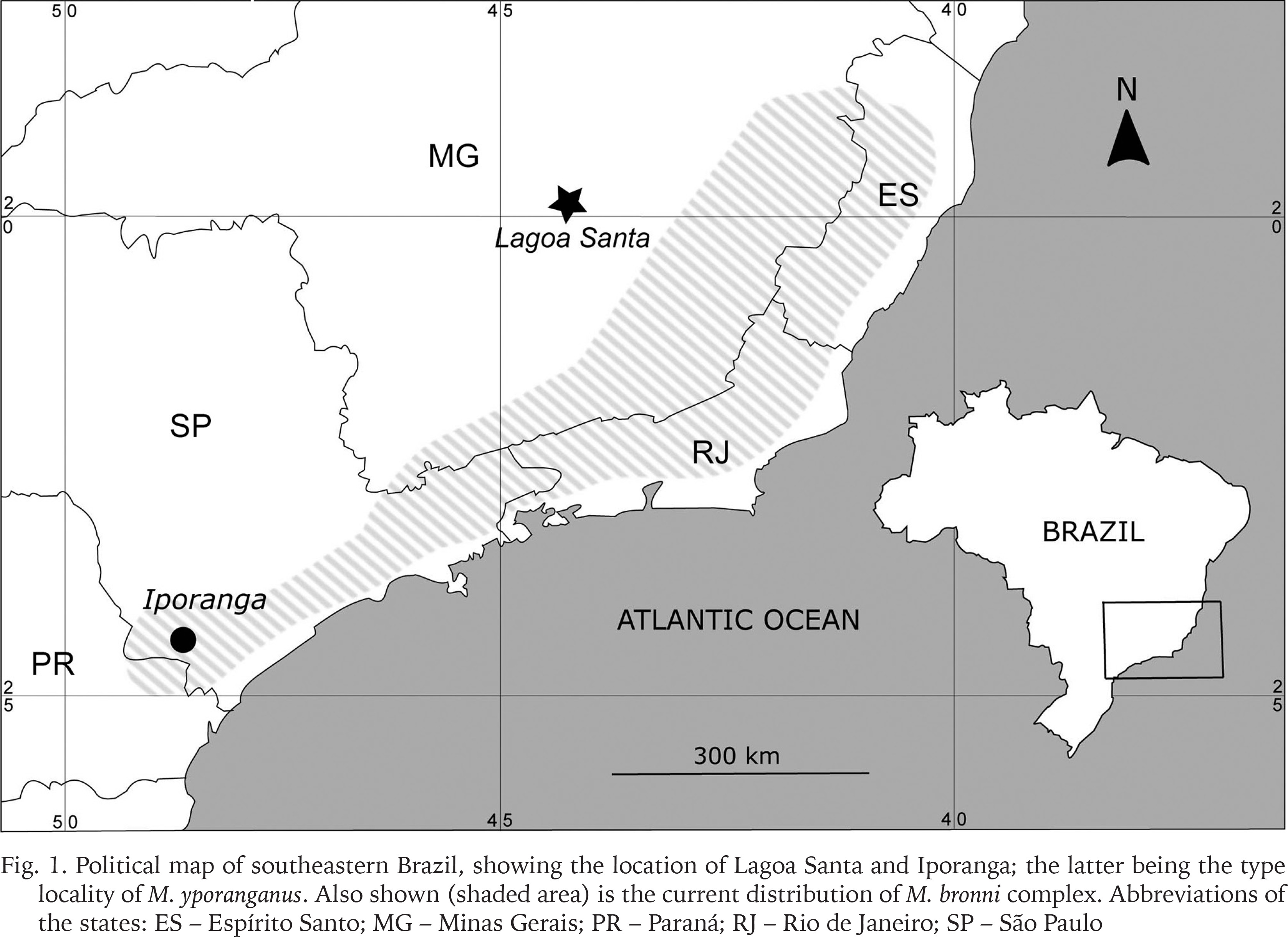
One such area is Lagoa Santa, in Minas Gerais state, southeastern Brazil, where shells of Megalobulimus have been preserved in an archaeological context in the region’s karstic caves. During the archaeological excavations of the Franco-Brazilian Mission (“Missão Franco-brasileira”) to Lagoa Santa in the 1970s, many megasnail shells were collected and, at the time, assigned to the species M. oblongus (O. F. Müller, 1774) and M. yporanganus (Ihering et Pilsbry, 1901) (Leme 1975b). However, the distribution of M. yporanganus (including subfossil material) is restricted to southern São Paulo state to northernmost Santa Catarina state (Fontenelle et al. 2019), and distant from Lagoa Santa.
A critical revision of that archaeological material and further specimens, allied with current taxonomic understanding of Megalobulimus in southeastern Brazil, indicated that the material from Lagoa Santa assigned to M. yporanganus belonged to a yet undescribed species. Herein, we describe it as Megalobulimus diluvianus sp. nov.
MATERIAL AND METHODS
ABBREVIATIONS
The following abbreviations are used throughout the text. Institutional acronyms: MHNJB – Museu de História Natural e Jardim Botânico da Universidade Federal de Minas Gerais (Belo Horizonte, MG, Brazil); MZSP – Museu de Zoologia da Universidade de São Paulo (São Paulo, SP, Brazil); NHMUK – Natural History Museum (London, UK). Shell measurements: Al – aperture length (distance across parietal region of aperture, from columellar lip insertion to palatal lip insertion); Aw – aperture width (distance, perpendicular to Al, from end of columellar fold on parietal region to the outer lip); D – shell depth (in lateral view, greatest measurement perpendicular to H); H – shell height (longest measurement parallel to columellar axis); Np – number of whorls of protoconch; Nt – total number of whorls; W – greatest shell width (in apertural view, greatest measurement perpendicular to H). Other abbreviations: coll. – collector(s); sh. – shells.
STUDY MATERIAL
The bulk of the material studied herein comes from the Franco-Brazilian Mission, which took place in 1971–1977. The gastropod shells collected were donated to the malacological collection of the MZSP. In total, the expedition collected 37 lots of shells, all from the Holocene site “Lapa Vermelha IV” in Confins municipality, in Lagoa Santa area.
We also analyzed additional material from Lagoa Santa, as well as from other caverns and archaeological sites in its surroundings, housed in the MZSP and MHNJB collections. Additional specimens belonging to the new species were found in the invertebrate paleontology collection of the NHMUK, collected in the first half of the 19th century by Peter Clausen (1801–1872) and acquired by the museum from him in 1844.
Further comparative material of Megalobulimus spp. was obtained from the MZSP collection for analysis. Measurements of the specimens were taken with a digital caliper (0.01 mm precision, rounded to the nearest 0.1 mm).
ARCHAEOLOGICAL SETTING
Lagoa Santa is situated in a karstic region in southeastern Brazil (Fig. 1). It covers an area of 356 km2 and contains not only the eponymous lagoon, but also rock shelters and limestone caves, being one of the most important archaeological sites in Brazil, related to the early occupation of the continent (dating back to ca. 12,000 BP; Araujo et al. 2012). It is considered the birthplace of Brazilian paleontology and archaeology thanks to the studies of Danish naturalist Peter W. Lund (1801–1880).
Fig. 1
Political map of southeastern Brazil, showing the location of Lagoa Santa and Iporanga; the latter being the type locality of M. yporanganus. Also shown (shaded area) is the current distribution of M. bronni complex. Abbreviations of the states: ES – Espírito Santo; MG – Minas Gerais; PR – Paraná; RJ – Rio de Janeiro; SP – São Paulo
Lund listed 149 extant and 32 extinct species of vertebrates from the region (formally describing several of them), but only mentioned molluscs in passing, for instance, referring to freshwater bivalves only to highlight the inundation of the caves during the rainy season (Lund 1836–1846). Likewise, Danish naturalist Peter Clausen listed 101 species of mammals between 1832 and 1840, but granted the mollusks only the brief comment “many terrestrial and fluviatile shells” (Clausen 1841). Luckily a few of his molluscan specimens reached the NHMUK. A greater focus on Lagoa Santa’s mollusks would come only with the Franco-Brazilian Mission (Leme 1975a, b) and later with the paleoenvironmental observations of Prous (1990, 2016).
The lots from the Franco-Brazilian Mission contained shells of M. oblongus and Thaumastus taunaisii (Férussac, 1822) (Orthalicoidea, Megaspiridae), both associated with firepits in Holocene strata, and M. yporanganus, in sediments from the Pleistocene/Holocene transition (Leme 1975b).
SYSTEMATICS
Superfamily Rhytidoidea
Family Strophocheilidae
Genus Megalobulimus K. Miller, 1878
Megalobulimus diluvianus sp. nov. (Figs 2–11)
Figs 2–11
Shells of Megalobulimus diluvianus sp. nov., with detail of the protoconch of selected specimens: 1–5 – holotype MZSP 161551, from Gruta da Lapinha (ventral (2), lateral (3), and dorsal (4) views, and protoconch detail (5)); 6–7 – paratype MHNJB 1654-18, from Lapa Vermelha IV (dorsal view (6) and protoconch detail (7)); 8 – paratype MZSP 20442, from Lapa Vermelha IV, ventral view; 9 – specimen NHMUK G25267; 10 – paratype MZSP 15.588-4, from Gruta da Lapinha, protoconch detail; 11 – paratype MZSP 15.588-1, Gruta da Lapinha, protoconch detail. Scale bar for shells 50 mm

Megalobulimus yporanganus: Leme1975b: 174.
Megalobulimus yporangus [sic]: Leme 1975a: 138; Prous 1990: 281; Prous 2016: 123.
urn:lsid:zoobank.org:act:5A4C2FC1-4953-457A-BD2D-8390DF493012
Type specimens. Holotype: MZSP 161551 (Leme & Papavero coll., 1963). Paratypes: Brazil, Minas Gerais state. Lagoa Santa municipality, Gruta da Lapinha [“Lapinha Cave”]: MZSP 15588 (6 sh., Leme & Papavero coll., 1963), MZSP 29305 (1 sh.). Confins municipality, Lapa Vermelha IV [“Vermelha IV Cave”]: MHNJB 1654-18 (1 sh.), MZSP 20442 (1 sh., Projeto Lagoa Santa, 1971), MZSP 39010 (1 sh., Projeto Lagoa Santa, 1971), MZSP 39003 (1 sh., Projeto Lagoa Santa, 1971). Matozinhos municipality, Lapa do Santo [“Santo Cave”], MZSP 133360, 1 sh., Gruta Cuvieri, MZSP 144003, 2 shs.
Additional material. Brazil, Minas Gerais state. Undetermined caves: NHMUK G1505 (1 sh., P. Clausen coll., purchased 1844), NHMUK G25266 (1 sh., P. Clausen coll., purchased 1844), NHMUK G25267 (1 sh., P. Clausen coll., purchased 1844).
Type locality. Brazil, Minas Gerais state, Lagoa Santa municipality, Gruta da Lapinha (“Lapinha Cave”; 19°33'43"S, 43°57'35"W, ~700 m elevation).
Etymology. Following old “stratigraphic” nomenclature from Europe, Lund (1836–1846) named the soil layer in Lagoa Santa containing extinct megafauna as “diluvium”. The specific epithet is a reference to that nomenclature.
Stratigraphy and age. Lund’s “diluvium”, a red layer lacking internal stratification and typically overlaid by calcite, was formed during the beginning of the Holocene under a more humid climate (Auler & Piló 2016). The new species disappears from the fossil record following Lund’s diluvium and still belongs to the early Holocene (Prous 2016).
Diagnosis. Shell medium-sized for genus, elliptical-elongated. Spire short, with narrow base (proportionately to body whorl). Nepionic ribs well-marked, flattened towards suture, becoming progressively more spaced before fading away. Body whorl tall and wide, with spiral malleations, lacking granulated microsculpture. Columella straight, long. Aperture elongated.
Description. Shell medium-sized for genus (mean H – 89 mm), elliptical-elongated (Figs 2, 6, 8, 9), lightly dorsoventrally compressed (D/W – 0.8; Fig. 3), with circa 5 whorls; imperforated to rimate. Whorl profile slightly convex. Suture shallow. Spire short, with narrow base (proportionately to body whorl). Protoconch (Figs 5, 7, 10, 11) of circa 3½ whorls; first 2½ whorls narrow and flattened; starting at 1½ whorl, sculptured by straight well-marked axial ribs, lightly prosocline, flattened towards the suture, regularly spaced; from 2½ whorl onwards, ribs become lightly curved, progressively more spaced, and then fade away. Penultimate whorl rapidly expanding into body whorl, with regular fine growth lines and radial granulated microsculpture. Body whorl tall and wide, with irregular growth lines and spiral malleations, lacking granulated microsculpture (Figs 4, 6). Columella straight, long, with wide but not much pronounced fold. Aperture elliptical, elongated, circa half shell height. Peristome only lightly thickened, reflexed forming a gutter-like structure external to the aperture. Periostracum not preserved.
Measures. Holotype: H – 93.0 mm, W – 52.0 mm, D – 42.0 mm, Al – 50.5 mm, Aw – 24.0 mm, Np – 3.4, Nt – 5.0; paratypes (n – 15): H – 89.0 mm [84.8–93.5], W – 49.3 mm [45.9–52.0], D – 40.0 mm [36.5–43.5], Al – 22.7 mm [20.0–24.5], Aw – 47.0 mm [40.0–50.5], Np – 3.5 [3.4–3.6], Nt – 5.0 [4.9–5.1].
Distribution and habitat. All specimens of Megalobulimus diluvianus sp. nov. are in a subfossil state, having been collected in rock shelters and limestone caves in the area of Lagoa Santa (Fig. 1). The specimens collected by Clausen lack precise locality but, in all likelihood, also stem from Lagoa Santa region. More detailed information is only available for the specimens collected in the site Lapa Vermelha IV, which stem from archaeological strata from the Pleistocene-Holocene transition, when the climate was colder and drier than at present (Leme 1975b). There are no known living representatives of this species, so the species is presumed extinct.
DISCUSSION
TAXONOMY
The flattened early protoconch with widely spaced ribs seen in Megalobulimus diluvianus sp. nov., as well as the wide and tall body whorl and elongated aperture, are some of the diagnostic characters of a group of Megalobulimus species from eastern Brazil known informally as the “M. bronni complex”, which includes six species (Bequaert 1948, Leme 1964): M. bronni (L. Pfeiffer, 1847), M. fragilior (Ihering, 1901), M. garbeanus Leme, 1964, M. klappenbachi Leme, 1964, M. parafragilior Leme et Indrusiak, 1990, and M. pergranulatus (Pilsbry, 1901). The species in the M. bronni complex are distributed along the mountain chains that follow the southeastern Brazilian coast (such as the Mantiqueira Mountains and the Serra do Mar in São Paulo and Rio de Janeiro states) onto the plateaus of Minas Gerais and Espírito Santo states (Fig. 1; Bequaert 1948, Vieira & Simone 1990, Oliveira & Almeida 2000, Simone 2006).
In comparison to the species of that complex, and in particular to M. bronni (Figs 12–13), M. diluvianus sp. nov. has a more elongated shell, sculptured by straighter and more spaced ribs, but lacking microsculpture on the body whorl. Its aperture is likewise more elongated, with a longer columellar region. M. diluvianus sp. nov. is most reminiscent of M. klappenbachi (Figs 16–17), from southern São Paulo state, sharing a similar spire and aperture profiles. It differs by the more regular and well-marked protoconch ribs, a larger aperture with a more rounded base, and the lack of a granular microsculpture on the body whorl. Furthermore, the spire of M. klappenbachi is proportionately much smaller in relation to the remainder of the shell.
Figs 12–17
Shells of Megalobulimus spp. for comparison, with detail of protoconch: 12–13 – M. bronni, specimen MZSP 92421, from Além Paraíba (MG), ventral view (12) and protoconch detail (13); 14–15 – M. yporanganus, holotype MZSP 64144, from Iporanga (SP), ventral view (14) and protoconch detail (15); 16–17 – M. klappenbachi, holotype MZSP 29317, from Iguape (SP), ventral view (16) and protoconch detail (17). Scale bar for shells 50 mm

As the present material was once assigned to M. yporanganus (Figs 14–15), a comparison with that species is warranted. M. diluvianus sp. nov. differs from it by the widely spaced and straighter protoconch ribs (flattened towards the suture), lack of microsculpture on the protoconch, a lower and more rounded spire apex, an overall more elongated shell, and a straighter and elongated columellar region.
PALEOECOLOGY
The occurrence of M. diluvianus sp. nov. in the caves in Lagoa Santa has an ambiguous interpretation. Live snails can be attracted to the limestone caves due to the abundance of calcium (Dallinger et al. 2001) or to use them as shelters during aestivation (Fontenelle & Miranda 2017). Likewise, the shells could have been brought passively (usually post-mortem) into the cave by water, such as during flooding events (e.g., Vieira & Simone 1990). Those two options indicate, respectively, an autochthonous or parautochthonous (the shells are reasonably well preserved) origin of the snails. However, Lagoa Santa is also an archaeological setting, which brings the human factor into the fold.
Gastropod shells have been identified in archaeological contexts around the world. The live animals and/or their empty shells have been collected by humans for numerous reasons: food, production of dyes, tools and ornaments, as well as for magico-religious purposes (Claassen 1998). As such, it has been suggested that the peoples who inhabited Lagoa Santa and its surroundings since 12,000 BP were actively collecting the snails and/or their shells (Moreira et al. 1991, Prous 1990, 2016, Araujo et al. 2012). Araujo et al. (2012) argued that consumption of Megalobulimus spp. occurred in Lagoa Santa, as well as in other localities and traditions in eastern Brazil, such as Itaparica and Umbu.
Given the possibility of human interference and the difficulty in defining whether the presence of a given species is natural in the area, paleoecological and paleoenvironmental interpretations using shells from archaeological settings must be approached with caution (Lubell 2004a, b).
In Lagoa Santa, Prous (1990) assigned the disappearance of M. diluvianus sp. nov. (identified as M. yporanganus in that work) from the early Holocene stratigraphic record of Lapa Vermelha IV to climatic alterations. The change in climate was supported by Neves & Piló (2008), who in their sedimentological analysis of Lagoa Santa identified peaks in humidity around 9,700 BP and 2,000 BP. As such, Prous (2016) inferred that due to the increase in temperature and humidity, M. diluvianus sp. nov. was locally substituted by M. oblongus, simultaneously with human occupation of the area. Accordingly, M. oblongus became the most common species in later Holocene strata (Prous 2016).
With the present evidence, it seems that M. diluvianus sp. nov. was not only locally replaced, but rather went extinct. It is impossible to tell whether the causes of this extinction are related to climatic change, competition with M. oblongus (and/or other species), human consumption, or any combination of those.